(本題に戻ります)問題はSNPs解析に戻ります。再掲しましょう。
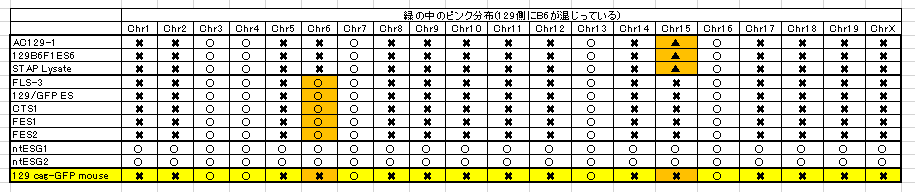
一番下のたまたま分析を受けた129 cag-GFP mouseの15番染色体のSNPsパターンは最上段3つの細胞とは違うというところを確認してください。これは若山さんの129マウスにマウスコンタミがあるからでしたね。解析を受けた129と上3つを作った時の129が少し違うのです。でも上三つは同じマウスからできている。そしてAC129はどうせ若山さんが「僕のマウス」ES-6を詰め替えたものです。しかし、STAP Lysateは違いますよね。これは小保方さんがChIP検査のためにかき集めた細胞だと証言していると伊藤さんが記者会見で語っています。人から取り戻している細胞ですね。このときに誰かが129B6F1 ES6を渡しましたね。AC129の実験時ではありません。小保方さんはF1のSTAP細胞として提出しているのです。
このことは何を意味するかというと、既に2012年の8月には小保方さんは嵌められているということです。そして原因は恐らく特許での対立です。若山さんはこの対立での経緯でこのまま放置できなくなった。万が一の時には小保方さんの捏造で逃げる準備が行われている。と考えると何となく辻褄はあう。でも、これはSNPs解析結果ですね。思い出してください。Nanogの発現解析はSTAP細胞とES細胞は違っていましたよね。伊藤さん。本当にSTAP Lysateが残っていましたか?
>>
ただ、唯一やったのが、あ、GRASの方に残っていたChIP seq のinput 、しかもSTAP細胞のインプットはDNAとして残っていたので、それが10ページバワーポイントの、えーと、ところに書いてある、3番のアイテマイズして書いてあるところですけれども、これに関しては、えーと、30倍になるだけ、他のゲノムと同量となるまで読んでいます。そして、読んだ結果、この、同一であるということを、ま、認定したということで、それ以外に、それ以外に関しては、ま、残っていませんのでやってない、やってません。(*ttps://www.youtube.com/watch?v=M9oJGioHvIQ 1:15:00から)
因みにここで伊藤さんはこのたくさん集めた細胞は別の実験だと言ってますね。つまり彼らはこれがAC129の実験時のSTAP細胞ではないことを聞いているんです。にもかかわらず、「公開データ再解析の結果に よれば、論文に記載された実験の中では Letter Fig.4 に使われた可能性が高く」と書いているのです。嘘ですね。彼らはここで小保方さんが若山さんの「僕のマウス」ESを使ったと主張しているのです。そのくせNanogの件は無視している。それならこの時のESは何なのか。
GRASに残されている記録を全部発表してもらいたい。若山さんのこの頃の実験ノートを出してもらいたい。どうして小保方さんばかりに訳の分からない因縁をつけているんでしょう。小保方さんは調査チームを信用して実験ノートを提出しているのに、彼らは全コピーをNHKに違法に流出させて犯人の検挙もしてないだろうが。告発すらしていない。犯罪を知ったら警察に届けるという公務員義務違反をしていて恬として恥じない。日本の知的レヴェルは明らかに下がってますね。こんなのが博士だと思うと情けないね。ははは。
さて、いよいよこの時の129/Svは何なのかという問題に入れるわけです。FLS3以下5つの細胞株の6番染色体が一番下の129 cag-GFP mouseとは違うことを確認してください。これも又別のコンタミマウスなんですね。でも他のB6のSNPsパターンは全部同じだということも確認してください。これって市販の129/Svマウスではありませんよね。アルイミオウジ氏も疑義を呈している。でももし129 cag-GFP mouseだったら18番染色体にヘテロでGFPが入っているはずです。
やっと最初の問いに戻りましたね。どうしてFES3のGFPは岡部マウス側のAcr-CAGのみが調べられて、母親側の18番にCAGが無いかどうかを確認されてないのか。もしそれがあったらFES1,2には無いはずのものですよね。仮にterで作ったという記憶が間違いでX1であったとして、しかもそれは市販の129/Svではなく、更に若山さんの129/Sv-CAGホモだったとして、更に、どうして7年も前のコンタミマウスのSNPsパターンの同じ親にぶち当たるのだ。
FES1,2ともに中身の細胞は若山さんによってFLS3に入れ替えられているということです。
さあ、大問題です。若山記者会見です。
*ttps://www.youtube.com/watch?v=hD3Wd9cpo78
FLS1-8は15番にヘテロで4コピーのGFPがあり18番には無い。「僕のマウス」シリーズには15番にはGFPは無く、18番にホモで1コピー入っている。誰でも知ってることですね。
ところで15番にGFPが入っているという第三者機関の発表は間違いとされた。遠藤さんが発見したことになっているが、それはともかくとしてではFLSの18番にはGFPが無いという結論はどうなっているのか。一方が間違っているのに他方は合っているというのも変な話しで、その場合は間違った原因を述べなければならず、又同時に18番を調べた手法の確認も必要です。でもそれはうやむやになっているままなんですね。SNPsパターンは129x1/Sv-CAGホモマウスの特徴がほとんど全染色体に分布してますよね。どうなってるんでしょうかね。
今、アルイミオウジ氏がいくら兄妹交配を重ねても決してホモ化しないSNPsがあるなどというトンデモ主張中ですが、又、テヘで終わるのでしょうかね。そんな現象があるのなら、そもそも報告書のSNPs解析は意味を持ちませんね。
有る特定の129マウスが近交系マウスになったというのは兄妹交配回数が20回を超えて、基本すべての対立遺伝子がホモに揃っているものです。129特異的SNPsというのはその全遺伝子の並びの中で野生型と比較してSNPとして違っている箇所が特定されているということです。近交系マウスの中では全部ホモになっているSNPsです。これは定義なので近交系マウスの中のヘテロなSNPsというのはあり得ないんです。まずヘテロなSNPsになっていたら近交系マウスではない。以前から説明せずに一人よがりする人ですから他所から見て何とも言えませんが、近交系マウスの中でヘテロなローカスというのはいくらでもある。全遺伝子がホモに揃っているというのは実験目的での実用の範囲で言うものです。実際にはヘテロなアレルは残っています。以下が参考になる。
*www.anim.med.kyoto-u.ac.jp/Kuramoto/contents/ExpAnimGenet_1_inbred.pdf
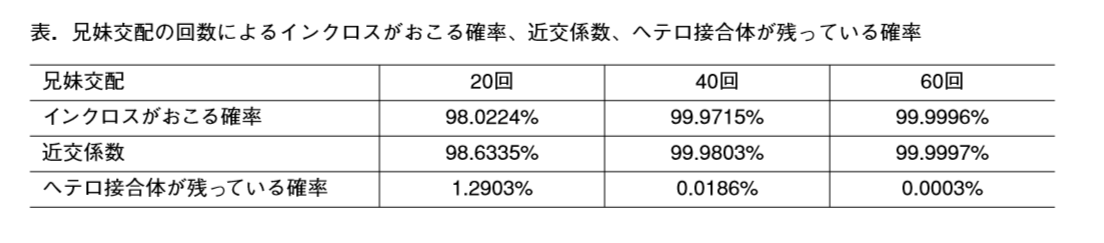
マウスの場合は胎児から成熟して妊娠できるようになるまで20日と50日の合計70日かかる。20回の交配というのは2.8年、40回の交配というのは7.7年、60回の交配は11.5年です。マウスの全塩基数は30億個程度です。60回やってヘテロ残存率が0.0003%になった時の数は9000個です。染色体数を20として一本に450か所ある。20回の交配後既に近交系マウスとして扱われるので、その場合では38,709,000か所残っているので染色体1本につき約2百万か所のヘテロなアレルがあることになる。
このヘテロなアレルは両親の違いがそのまま残っているだけのものです。これを更に兄妹交配して世代を重ねていくとヘテロだったものがどちらかに固定されていくんです。一旦固定されると何度交配してもホモなんですから子供には同じ核酸が分配される。これがインクロスと呼ばれる交配なんです。全部がインクロスになったら完全クローンになりますが、理論的にはこのやり方では無限に繰り返してもヘテロはわずかに残ってゼロに限りなく近づくが完全なゼロにはならない。
この問題と、ある近交系マウスに特異的なSNPsというのは無関係です。これは野生型と違う核酸になっている場所のことで、近交系マウスではほぼすべてのアレルがホモですから、ホモの中で野生型と違っている場所という意味です。ヘテロなSNPsなんて子供を造ったら分かれてしまうのでその近交系マウスに特異的SNPということにならないんです。定義の問題ですね。ホモで且つ野生型でない場所。そして近交系マウスの種類によって核酸の種類も場所も違っているものを特異的SNPsという。
まあ相変わらず一人で分かった気になってまともな説明が無いので端からは何とも言えませんが、この問題の解にはなってない筈です。
アルイミオウジ説の話になったので序に書いて置きますが、彼はntESG1,G2のGをgenerationと解して、最初のntESがG1で、それを使ってキメラマウスを作って更にntES化したのがG2と推測している。そして実際に若山さんはこのやり方で20世まで作っていて別の論文があるわけです。でも太田さんのG1ははっきりしていて、2008年論文で2005年論文で作ったF1のntESを解凍して使った。その解凍した細胞を129B6F1G1と書いているんです。すると解凍する前の状態のラベルはどう書いてあるのかということになる。株分けした時に以前の分に129B6F1G2と書いたとしたらこれはgenerationの意味ではありません。ただ、日経サイエンスに太田さんは自分は2種類の受精卵ESと4種類のntESを作ったと証言している。太田さんに聞けばわかることで、彼には説明義務がありますよね。でも、チューブのラベルと中身が違っていることの説明は彼一人の判断では出来そうもありませんよね。
(残された問題の整理)さて、問題の数も少なくなってきましたね。ただし、解の困難さは増してきました。ここらで残されている問題を整理しておきましょう。
①FLSは「僕のマウス」を渡したと一方的に証拠の提出もなく若山さんが主張しているが、実際の検査で出て来たのはAcr-CAGのB6(岡部マウス)と市販のGFP無しの129X1/SvのF1だという報告書の調査結果となっている。しかし、129側の18番染色体に1コピーのCAG-GFPが入っているのではないかという疑義がでた。なぜなら、SNPs解析結果がこの129は「僕のマウス」の片親であることを示唆しているからだ。最初に放医研がFLSを検査した時、15番にヘテロに4コピーのCAGがあり、18番にはGFPが無かったと発表していたが、後に15番のGFPが否定されたが18番に関しては何も触れられなかった。又間違いの原因もちゃんと説明されていない。内在性アクロシン遺伝子を間違えたのならホモだと思うはずでヘテロに間違うというのは考えにくい。調査自体がとてもあやしい。すると18番に無かったというのは本当なのかという疑義が残るということです。理研のチームはGFPを自分でちゃんと確認したかということです。この説明も無い。
②小保方さんはなぜローザだという説明を受けたのか。彼女は徒手空拳で何の理由もなくローザと書く動機が無い。誰かがローザだといった筈である。
③なぜカルスキメラ子の8番に3番染色体欠失があるのにAcr-GFPがないのか。多分これが我々素人にとっては最大の難問になりそうですね。というのも他のことは誰かが何かをしたとか、し忘れたとかいう可能性を思いつけるが、この問題だけはど素人ではなぜこうなるのかの一つの可能性すら思いつけない。
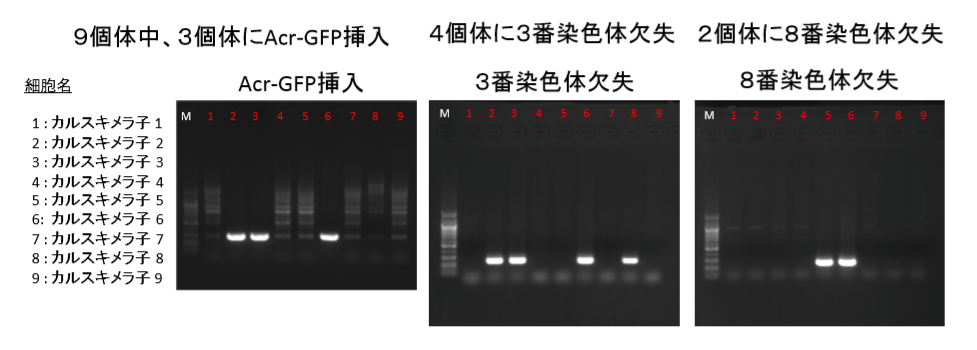
欠失とAcr-GFPはB6の同じ3番染色体上にある。一つの細胞に欠失が起きてそれが分裂して増える。最初は一つの細胞です。ここにAcr-CAGがあったらその後の体細胞分裂でGFPはずっと引き継がれる。欠失は同じなのにGFPが無いということはあり得ないんで、その場合は欠失の種類が違うということしか考えられない。逆にGFPありとGFP無しの細胞に同じ欠失が起こることも無いですね。まあ、せいぜい突然変異でGFPが消えたというなら分かりますけどね。そんな確率で起きることですかね。
この表ではB6に129がコンタミしている場所を指摘していますが、129にB6がコンタミしている場所を提示していませんね。もう一段深めて推定する発想力が無いんですね。そういう発想力があったらマウスコンタミの無いB6と129のデータをコントロールとして並べますよね。客観的たろうという姿勢が無いんです。まあ、科学者としての資質としては相当低い人々でしょうね。一般の会社に入っても役だたずでしょう。こういう奴って悪いことばっかり考えるんだよなあ。ときどき見かけるタイプ。流出させてはいけないデータを羽織ゴロツキとかNHKなんかに流す。
NHKに調査チームが病床の小保方さんの手元から奪い取るように持ち去った実験ノート3冊分の全コピーが流出するという犯罪があった。誰が流出させたのかをNHKが答える義務はない。又理研内での自己点検報告の草稿が作られたとき、それが毎日新聞の記者にリークされた。このリーク者を毎日新聞が公表する義務はない。しかし、二つの公務員としての犯罪が行われたことが明確であるにも関わらず理研がこの事件を警察にとどけることが無かった。これはこれ自体が犯罪ですね。
これは追及されるべき事柄です。日本は法治国家では無いのか。
内部告発も取材源秘匿の自由もちゃんと保証されている。でも違法行為は警察が取り締まるという義務も又課せられているんだとということを忘れてもらっては困る。理研は今からでもNHKに小保方さんの実験ノートを流出させた犯人を捕まえるよう依頼しないといけない法律になっている。無論、その際でも、NHKは警察に対して犯人が誰かを教えなければならないというような義務はありません。取材源の秘匿は法で保護されている。ただし、裁判所の令状があると別ですけどね。裁判所は裁判所で個々のケースで令状を出すか否かは判断します。このケースでは出ませんね。でも警察はそんなもの無くても犯人をすぐ特定しますよ。捜査権限がありますからね。実験ノートに関与した者なんて限られたわずかな人数です。こんなことをそのような人間が調査したとするサンプルの中身に工作しなかったなどと考える方がどうかしてますよね。何れ、その問題は最後の課題となるでしょう。
(①の問題から順番に)SNPs解析結果は129が「僕のマウス」の片割れだと示唆しているにも拘わらず、その18番染色体にCAG-GFPが無いという結論は放医研の出したときのままになっている。そしてB6側の結論が間違っていたと判明した後に理研が129側を確認したか否かも知らされていない。
放医研の調査結果を若山さんが記者会見発表して後に、遠藤氏がそのデータを若山さんから貰って、放医研の出した15番にヘテロに4コピーのCAG-GFPがあると発表した結論は間違いであったと言った。放医研はCAG-GFPを探していたので、CAG-GFPのプロモーターの位置をプライマーで挟んで探していた。その時に、そのプライマーではアクロシン遺伝子の有る場所を検知してしまったのだという。支離滅裂の説明ですね。意味が分からん。

まず不可思議なのはFLSの15番に4コピー入っているとしたPCR産物は何かということです。どういうプライマーで挟んだのかということです。CAG-GFPの挿入された場所を探している。CAG人工遺伝子の並びそのものをプライマーにかけるということになる。
若山さんは自分のマウスはCAGだと説明していますね。だから放医研はCAGを探そうとした。そして順序はともかくとして「僕のマウス」には18番にホモでCAGがあるとした。若山さんは今まで自分のマウスのGFPがどこに入っているのか調べたことがなかったのでしょうかね。岡部マウスはもらった時に3番に入っていると聞いていたわけでしょ。岡部マウスは放医研では調べていませんよね。でも記者会見では3番だと答えている。普通自分のマウスのGFPがどこに入っているかは調べるよね。そもそもマウスの飼育はときどきコンタミが無いかどうかのSNPs検査にも出すものですよね。

FLS1-8の18番にはCAG-GFPが無いが、「僕のマウス」ESにはある。しかもホモで入っていると。
では、15番はどうか。FLS1-8にはある。しかも「僕のマウス」ESには無い。ところがこの15番は間違いだったという。では18番は正しいのか?
これが分からないわけです。桂報告書もGFPが何番染色体に入っているかをリスト上に記載していない。15番を間違えるような奴が18番は間違えなかったとどうしていえるでしょうか。そもそも探したプライマーを明らかにしていない。本当に両方とも同じプライマーで挟んだのかということすら疑われる。18番のCAGが正しいなら15番のCAGも正しいはずだ。アクロシン遺伝子はヘテロに4コピーなんてあり方はしないぞ。これは別のプライマーじゃないのか。
記者会見の画像ではプライマーのところが写されてないものもあって分かりにくいので、片瀬久美子氏のブログのを参考に以下に貼り付けて置こう。
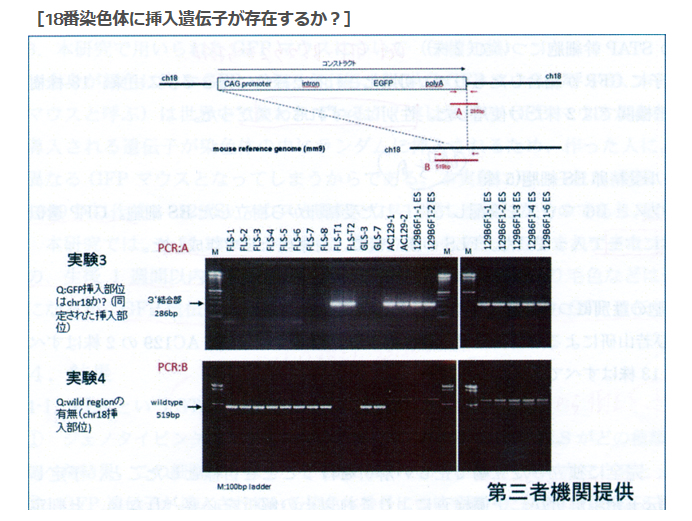
上にどういうプライマーで挟んだのかが図示されている。コンストラクトというのは人工遺伝子のことで、無論これは若山さんがロックフェラー大学でB6にヴィルスヴェクターを使って遺伝子挿入したときのコンストラクトです。若山さんが行ったのですからその遺伝子構成は若山さんが知っていて、これを探すときに若山さんは放医研の知り合いに教えている。で、その末端につけられているポリAのあるあたりの部分にプライマーを設定した。要するに若山さんの人工遺伝子の尻尾を探したわけです。
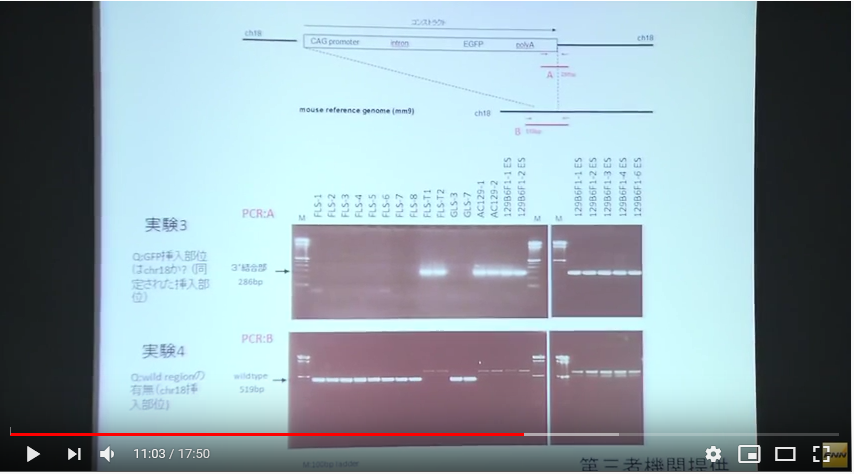
実験3は18番染色体を調べた結果FLS1-8には存在せず、所謂「僕のマウス」シリーズにはあった。従って、若山さんの「僕のマウス」のGFP人工遺伝子は18番に入っていたことが分かったというものです。
このプライマーが分からないわけです。上の図のAの長さは286bpと書かれていて、プライマーの長さは通常20bp程度を前後に二か所設定するわけです。前はポリAのある場所ですが、後ろはコンストラクトの外に表示されている。でも、これは人工遺伝子全体が挿入された場所が分かっていませんからこの後ろのプライマーのDNAの並びは分からない筈ですね。プライマーが分かっていたら既にどこに有るかわかってるということですから調査する必要はない。
ど素人が常識的に若山さんの人工遺伝子を探そうとするとき、コンストラクト全体の遺伝子の並びは分かっていますから、頭の20bpと尻尾の20bpで挟みますね。そして染色体ごとに探すとまずGFPの存在している染色体番号は判明しますよね。もっとも頭の並びはプロモーターの並びなので内在性のプロモーター位置を掴んでしまうかもしれませんが、後ろの20bp以上並ぶポリAはDNAにはないので特定できそうですし、それも何らかの紛らわしいことがあるのなら、中にあるEGFP遺伝子の頭と尻尾に設定してもいいですね。要するに何番染色体に存在しているかということはそんなにむつかしくなさそうに思われる。
ところがこの実験3は妙なプライマー設定だと見受けられる。後ろのプライマーはコンストラクトの外に表示されてませんか。これって挿入位置が既に正確に分かっているコンストラクトの存在を再確認しているだけではないのか。
また、実験4は何をしようとしているのかがど素人には分からない。こちらは全体の長さが519bpです。そして図を見る限りはコンストラクトの少し前の位置から後ろは実験3のプライマーの後ろと同じようです。つまりこの位置の中にコンストラクトがあることがあらかじめ分かっているということです。そしてその区間にFLS1-8ではワイルドタイプの遺伝子が並んでいるのだということなのでしょうが、では「僕のマウス」シリーズではどうかというとバンドが何もない。つまりその中にコンストラクトが挿入されて長くなっているから519bp以上の長さになっているからこのバンドのラインには出て来ないんですね。
これって、どこに入っているかは事前に正確に分かっているということじゃないですか。若山さんは挿入位置を教えて、それがあるか無いかを放医研に確認させただけではないですか。
となると、岡部マウスの情報も分かった上で確認させているのではないかと疑われて来る。
岡部マウスは3番に入っていることは放医研で調べたことにはなってなくて、かつ記者会見では3番と若山さんが言ってますから、あらかじめ岡部さんから教えられている。でも、この調査ではアクロシンなんて思いもよらないという前提で放医研にCAG-GFPの在処を探させているということになっている。
マウスの15番には内在性のアクロシン遺伝子が存在しています。こちらは実験3,4の後です。FLSには18番にGFPが無かった。でも蛍光してますからどこかにはある。で染色体を虱潰しに調べたら15番で引っかかってきたという。変ですよね。これはCAGの遺伝子配列をプライマーに使っている。若山さんのコンストラクト全体の中でプライマー設定されている。15番にはEGFPはありません。因みにGFPとEGFPは同じです。蛍光発色の強化されたものです。
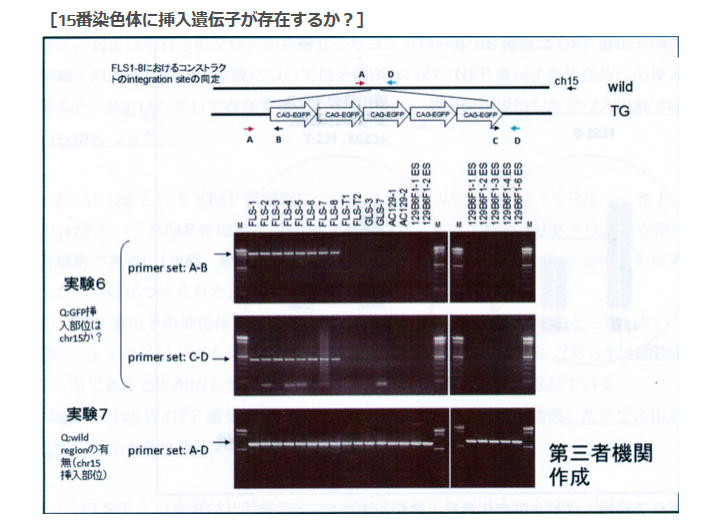
実験6は15番を調べたものと若山さんは記者会見で解説した。このPCR結果はあり得ないものです。GLSに無いのは当たり前ですが、そもそも15番にはGFPはない。それがFLS1-8には出ている。これは何なんでしょう。
そして実験7はA-Dで挟んでいる。これって若山さんのコンストラクトの中です。絶対に出ません。何なんでしょうか。
ひょっとして岡部マウスのコンストラクトを使ってないかと考えてみるとどうなりますかね。頭にアクロシンプロモーターがついている。すると15番にある内在性アクロシン遺伝子のプロモーターを掴むということはあるんですね。そして後ろのDの並びがあったかどうかでしょうね。
(どうしてこんなに分かりにくいのか)まずは実験3と4から確認していきましょう。見やすいように再掲します。
プライマーAはコンストラクトの後ろの方にポリAの並びがあって、その辺りからコンストラクトの末尾の少し後ろで挟んでいる。赤い矢印がそれですが、その長さは286bpと記載されている。そしてプライマーBはコンストラクトの少し前からプライマーAの後ろの位置と同じく、コンストラクトの少し後ろで挟んでいて、その長さは519bpと書かれていて、割り算するとプライマーAの長さは実寸でプライマーBの55%の長さです。この図でも赤の矢印とバーの長さと数値はそれを正しく示している。でも私は最初プライマーAはポリAの付近を挟んでいると思いましたが、他の人はどうなんでしょうかね。プライマーAのバーの長さと上の[CAG promoter---intron---EGFP---polyA]の概念図と比較すると尻尾の部分と誤解しますね。でもプライマーBはコンストラクトの長さを縮小していますよね。破線で縮小が示されている。プライマーの長さは数値で明確に書かれていますから、プライマーAはEGFPまで含んでいるんですね。全体の55% です。どうしてこんな紛らわしい表示にするのでしょうかね。全体を縮小しなかったらいいだけですね。何も面倒でないどころか、その方がシンプルで分かりやすい。縮小している意味が分かりませんよね。人が嘘をつこうとするときこういう姑息な手段を取りがちだということはまあ、警察のようにそれを仕事にしている人々だとか、そうでなくても比較的長く生きて経験豊富な人たちにとってはお見通しでしょうかね。しなくていいことをしている。分かりにくくするため以外には考えられない操作ですね。
コンストラクトの少しでも外側を使わなかったら実験4は出来ないというのは当たり前ですが、少し外側がプライマーとして知られているというのは事前にどこに自分のGFPが入っているかを知っているということです。
プライマーはAGCTの4種の塩基の並びの20桁程度を使います。4の20乗ですから組み合わせ数がほぼ1兆1千億程度になる。その20桁と同じ並びが他の場所にある確率は1兆1千億分の1ということです。マウスの全DNAの桁数は30億塩基程度ですから、余裕で同じ場所は無いと言っていいわけです。
[18番染色体に挿入遺伝子は存在するか?]ということを示している若山さんの記者会見での説明資料にある実験3は二つある前後のプライマーの後ろ側は若山さんのコンストラクトの外側にある。つまり挿入遺伝子は18番に有るということはあらかじめ分かっていて、かつ、どの場所にあるかまで正確にわかっていて、その確認をしているPCR実験です。もし場所が分かっていなくて分かっているのが若山さんのコンストラクトの遺伝子配列情報だけだったとしたら、その外側のプライマーはコンストラクト全体がどの染色体のどの場所に入ったのか分からないのですから作れません。
この実験はすべてが分かった後にその確認のために行われた実験だということになるわけです。[18番染色体に挿入遺伝子は存在するか?]という表題も又ミスリーディングなもので、このPCRで18番に有ることを探し当てたのではありません。既にどこに有るかわかっているものを確認しただけです。
実験4はその明らかな証拠で、プライマーBは若山さんのコントラストを内側に含んだ場所に設定してある。プライマーは業者に依頼して作ってもらうわけですが、どこに入っているか分かって無いと頼めません。これはプライマーの内側にワイルドタイプの遺伝子配列があるものがバンドに出ているものです。若山さんのコンストラクトが入っていたらその分長くなりますから、このバンド位置には出ないということです。これも分かったことを確認しているのであって、記者会見の席上で放医研とやらに調べてもらったなんて大げさなものではありませんね。このくらいは自分のところでできますね。どこに有るか分からないときは業種や専門機関に調べてもらう。高価な機械が必要です。この実験3と4はそのようなものでなく、放医研に頼んだとしたら結論が出た後の確認実験に過ぎません。或いはそもそも自分のマウスのGFP挿入位置なんて作成初期に専門業者に出して調べてもらっているはずのものなのですから、情報を全部与えてただ確認実験だけを第三者の立場だと称して行ってもらっただけのものかもしれない。そもそも放医研は研究所としては正式に依頼を受けたことは無いと言ってる。では依頼されたのは誰なんでしょうね。
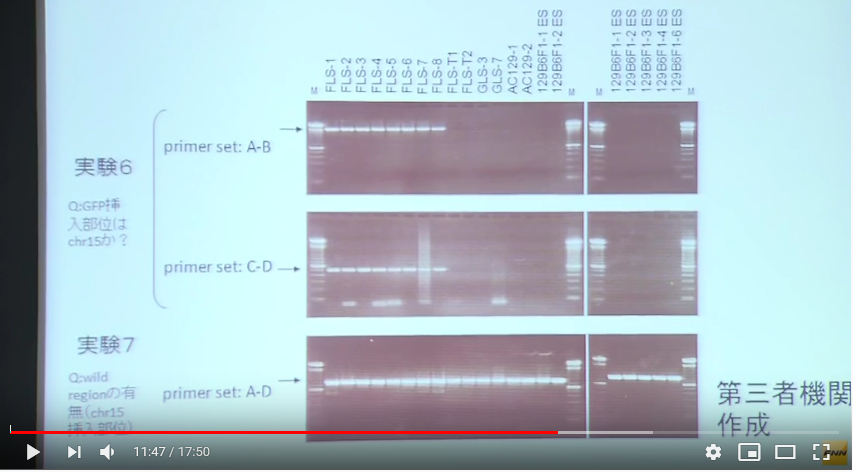
次は実験6,7です。これも見やすいように再掲します。
この説明図を見て異様に感じるのは実験3、4では若山さんのコンストラクトを探している、もしくは確認しているはずなのに、ここではそのコンストラクトが示されていないということです。その代わりにCAG-GFPが5つ繋がった概念図が示されている。若山さんのコンストラクトの中のCAG-GFPは1つです。何を探しているのか、又、何を確認しているのでしょうかね。
[18番染色体に挿入遺伝子は存在するか?]というタイトルの実験は実質的には18番染色体のどの位置に有るかということまでが分かっている上での確認実験でした。ところがこの実験は違いますね。[15番染色体に挿入遺伝子は存在するか?]というタイトルで、こちらは本当にあるかないかも分かってない。探しているのは若山さんのコンストラクトではありませんね。検査されているサンプルのGFPがどこにあるのかを探している。若山さんのコンストラクトは「僕のマウス」ESの18番にあったがFLSの18番には無かった。しかし、常時蛍光はしているからどこかに何らかのCAG-GFPは入っている。ではどこにあるのかを調べようとしているんですが、CAG-GFPを探しているのですからCAG-GFPの構造の前後をプライマーに取ればいいんですよね。CAG-GFPの構造は分かっていますからその中でプライマー設定するとそれがPCRにかかってきますからCAGがあるということは分かる。でもこれではどこにあるのかは分からない。そもそも常時蛍光しているんですから有ることは分かっていますからそんな検査は無駄です。これを知るにはある一定区分で区切ったたくさんの種類のプライマーを作って、それをワイルドタイプと比較すればいいとど素人でも思いつきますね。GFPが挿入されている断片は長いので上に出てワイルドタイプは下に出る。二本出たらそこにある。ゲノムは全部読まれていて並びは分かっていますからあとは更に厳密に範囲を狭めて行ったらいいですね。
ゲノムウォークという手法があって、挿入遺伝子の構造は分かっているのですからどちらでも片方のプライマーは設計できるわけです。他方がどこに有るかわかってないんですからここにアダプターを付けて取り敢えずの場所の検討をつけた後にそのアダプターに反応した場所を新たなプライマーとする方法がある。以下はタカラバイオのゲノムウォーキングキッドの宣伝の中の図です。
ど素人にはちょっとややこし過ぎて理解できませんが、まあ結論的に言えば場所の特定手法はちゃんとあるということです。業者に頼めば有料でやってもくれる。
ところが、放医研の第三者とやらは間違えたのです。マウスの15番染色体にはCAGは入っていなかったんです。ではこの15番染色体にヘテロで4コピーあったと報告したCAG-GFPは何であったのか。誰でも疑問に思いますね。竹市さんの疑問を貼り付けておきましょう。
>>
2014年7月22 日に訂正版を発表しました。
*ttp://www.riken.jp/pr/topics/2014/20140616_2/
2014年6月16日
CDB に保全されている STAP 関連細胞株に関する検証について
発生・再生科学総合研究センター
センター長 竹市雅俊
背景
STAP 論文では、脾臓から採取された免疫細胞が弱酸性にさらされることにより、多能
性を獲得することを報告した。STAP 細胞から増殖可能な細胞株として STAP 幹細胞が作
製された。STAP 論文に関する疑惑が明らかになった後、山梨大学若山教授は第三者研
究機関に STAP 幹細胞株の DNA 解析を依頼した。その情報は理研と共有され、理研は CDB
に保全されている STAP 関連幹細胞株の解析を進めてきた。
CDB 小保方研 STAP 幹細胞株の検討目的
CDB に保全されている STAP 関連細胞株(STAP 幹細胞、FI-幹細胞株)およびそれらから
作出されたキメラマウスの遺伝子情報を比較解析し、各細胞株間の遺伝子レベルの相違
と起源に関する客観的に検証可能なデータを得ることを目的とする。
CDB で保全されている小保方研細胞株の解析と結果
小保方研細胞株サンプルの遺伝子解析(遺伝的背景および CAG-GFP 挿入部位の確認)
• 小保方研細胞株サンプルのSTAP幹細胞6株に関して、遺伝的背景を6種のSNPマー カーを用いて検査したところ、以下の結果が得られた。GLS-1およびGLS-11につい ては核型の解析も行った。 1. FLS-3およびFLS-4: B6129F1: CAG-GFP, ♂ 2. GLS-1およびGLS-11: B6: oct4-GFP, ♂(核型の解析ではY染色体の一部に欠 失が見られる) 3. AC129-1およびAC129-2: 129B6F1: CAG-GFP ♂ (129 CAG-GFP マウス由来と されたが, 129B6F1由来であることが判明)。
• CDB では、若山氏から、STAP 幹細胞の CAG-GFP 遺伝子挿入位置の情報提供を受け、 上記 STAP 幹細胞株の CAG-GFP 遺伝子挿入部位を検証した。
STAP 幹細胞株 AC129-1 および AC129-2 は、18番染色体 GFP の挿入を持つ若山研 GFP マウスと同じ部位に、 1 コピーの CAG-GFP 遺伝子の挿入を持つことが判明した。かつ、相同染色体の両方に挿入されていることも若山研 GPF マウスと一致した。他方、FLS-3 および FLS-4 に関しては、15番染色体の片方の染色体に GFP 遺伝子が挿入されていることが判明した。また、CAG-GFP 遺伝子は複数コピーがタンデムに並んだ形で挿入されていた。これらの結果は若山研のサンプルの解析結果と一致した。
解析結果に対する見解
1.若山氏が提供されたとされる光るマウス(CAG-GFP 遺伝子保持マウス)から小保方氏が STAP 細胞を作成し、それを若山氏が受け取って樹立した STAP 幹細胞株に関して、保管されていたストックの解析から、CAG-GFP 遺伝子の挿入状況の違いにより、STAP 幹細胞は2種類の異なる遺伝子型のマウス由来であることがわかった。一方は、若山氏が提供した(CAG-GFP を18番染色体にホモで持つ)もの、もう一方は由来不明(CAG-GFP を15番染色体にヘテロで多コピー持つ)のものであった。
2.CAG-GFP を15番染色体にヘテロで持つマウスがどこ由来なのか、そのマウス個体が STAP 細胞から STAP 幹細胞が樹立された時期に若山研(あるいは小保方研)に生存個体として存在していたのかは不明であり、今後、さらなる検証を進める 誰でも15番にどうしてそんなものが見つかったのかを不思議に思いますよね。ここにはCAGはありません。にも関わらずどうして放医研の第三者といわれる人はここにCAGを4コピー見つけたのか。マウスの15番には本来の内在性アクロシン遺伝子があって、そのプロモーター領域もあります。仮に若山さんが岡部マウスのコンストラクトを聞いていてそれを教えていたのだとしましょう。ゲノムウォークするときに既知のプライマーがアクロシンプロモーターになっていた時には15番の内在性遺伝子領域をひっかけた可能性がありますね。でも、ここには4コピーなんて入っていません。4コピーあるのはCAGで3番に入っているのが岡部マウスです。間違えたにしても話がおかしすぎますよね。
(アクロシンプロモーター)放医研の第三者とやらはあくまでもCAGを探していたのだとしましょう。ゲノムウォーキング手法を使うとCAGが5個タンデムに並んだ断片が見つかったのだとしましょう。そして周辺を調べたらそこにアクロシンプロモーターが見つかったので15番だと思ったということですね。ところがそのアクロシンプロモーターは岡部マウスの人工遺伝子につけられていたアクロシンプロモーターだったということですね。
そして後に遠藤氏がそれに気が付いたという話になっているわけです。我々は誰が犯人か知っている。するとこのストーリーが作られたものだということはすぐわかる。その証拠が無いかということですね。
アクロシンマウスは若山さんは自分で維持飼育してますからね。「僕のマウス」B6でなければ、GOFのB6か、岡部マウスB6に決まっている。遠藤さんなんて出てこなくても可能性の中にはすぐ上がってくるものです。そもそも話が白々しいんですよね。あたかも自分は岡部マウスなんて考えもしてなかったなんて、最初のテラトーマからアクロシンが出でいて、このマウスが「僕のマウス」でないことは桂報告書の指摘するところです。FLSの時に「僕のマウス」を渡したということに実験ノートの証拠すらない。
岡部マウスの特徴は以下です。
>>
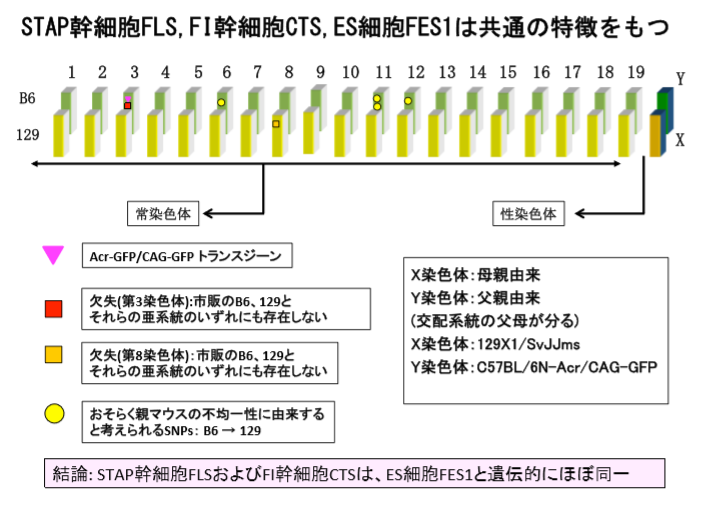
1)Acr-GFP/CAG-GFP 共挿入の位置、コピー数、周囲の塩基配列 表:STAP 幹細胞株一覧に挙げた 12 種類の幹細胞から STAP 幹細胞 FLS-T を除く 11 種 類の幹細胞株、それらの幹細胞が作製された 129 系統、および C57BL/6 系統の NGS によ る全ゲノム解析を行なった。その結果、Acr-GFP/CAG-GFP の共挿入は、STAP 幹細胞 FLS3、 FI 幹細胞 CTS1、そして ES 細胞 FES1 並びに FES2、ntES1 並びに ntES2, および 129/GFP ES の 7 株の第 3 染色体の同一部位に共通に存在することが判明した。また、Acr-GFP が第 3 染色体の片方にのみ挿入されていること(FISH により確認)、Acr-プロモーターの コピー数がどれも約 20 コピーであること、GFP 挿入部位を挟んで第 3 染色体の約 20kb の重複があることと、GFP 挿入部位に隣接して第 4 染色体 20kb 断片の逆向きの挿入があ ることも共通していることが判明した。これらの特徴は、2003 年に CDB 若山研が大阪大 学岡部研より導入した Acr-GFP/CAG-GFP マウスの特徴と完全に一致する。 (4P) ①3番染色体にAcr-GFP/CAG-GFP 共挿入がある。
②アクロシンプロモーターのコピー数は20コピー程度である。
③GFP挿入部位を挟んで約20kbの重複がある。
④GFP 挿入部位に隣接して第 4 染色体 20kb 断片の逆向きの挿入があ る。
⑤この特徴を共有しているのは2003年に岡部研より導入した岡部マウス、FLS3、CTS1、FES1、FES2、ntES-G1、ntES-G2、129/GFP ESである。
無論岡部マウスはホモ、その他はF1なのでヘテロに入っている。
ところで3番染色体には③④の二つの異常がある。しかし以下には欠失だと書かれていて、これが何なのかは上記引用箇所だけでは分からない。何度でも貼り付けましょう。
赤の四角は欠失と書かれていますよね。③④の中腹や交叉乗り換えとは違います。これって何でしょう。
>>
4)第 3 染色体と第 8 染色体の欠失変異
STAP関連11細胞株の全ゲノム解析から、第3染色体の5kbの欠失と第8染色体の17kb の欠失(第8染色体は129系統由来;第3染色体はB6系統由来)が上記STAP幹細胞FLS3、 FI 幹細胞 CTS1、および、ES 細胞 FES1 並びに 129/GFP ES だけに共通に存在することが 判明した。この2箇所の欠失は、STAP 幹細胞 FLS および FI 幹細胞 CTS の全ての株にも 共通に存在することがPCR産物の塩基配列決定により確認された。一方、この両欠失は、 市販の 129 の亜系である 129 x 1/SVJJmsSlc(SLC)と 129+Ter/SvJcl(CLEA)のいずれにも存在しない。また、この第 3 染色体の 5kb の欠失も、市販の B6 の亜系である C57BL/6JJmsSlc (SLC)、C57BL/6NCrSlc (SLC)、C57BL/6J (Charles River)、C57BL/6NCrl (Charles River)、C57BL/6JJcl (CLEA)、C57BL/6NJcl (CLEA)のいずれにも存在しない。 さらに、2010 年に若山研で受精卵凍結された Acr-GFP/CAG-GFP マウスにも存在しなかっ た。 もし、これらの細胞が論文に示されていた(129 x C57BL/6)F1 から作製された株で あるなら、これら 2 個所の欠失の両方、または片方が市販の 129 系統、C57BL/6 系統の いずれかに存在していなければならず、STAP 研究の行なわれた 2 年強という期間でこれ ら 2 個所の欠失が生ずることは考えにくい。従って、この結果は、これら 4 種類の細胞 が、論文に示されていた(129 x C57BL/6)F1 マウスから直接作製された株ではないこ とを明確に示している。(6P)8番は129側ですので今は論じない。B6の3番に別の欠失5kbがあるんですね。そしてそれを共有しているのは今度は、FLS3、CTS1、FES1、129/GFP ESです。岡部マウス、FES2、ntES-G1、ntES-G2には無い。
さて、いよいよ謎のキメラ子です。もう一度貼り付けます。
- 2019/11/07(木) 18:02:54|
- AC129
-
-
| コメント:3













