『一枚報告補記2』(基本にある虚偽)小保方さんに太田ESの細胞塊を渡されて、それが普段と違うということに気づかなかった若山さんは、最初のキメラ実験のときにそれをナイフで20個ほどの大きさにカットして胚盤胞に挿入したんですね。ES細胞の大きさは以下の図の右にある。この細胞の20個分は左の写真にはなりませんよね。そんな大きなパイプは胚盤胞に挿入できません。左の写真は実際に小保方さんが渡した細胞塊をナイフカットして挿入している時の写真です。中に入っているのはES細胞ではありませんね。
若山さんは以後ずっとES細胞で騙されてキメラを作り続けていたと言ってることになる。嘘ですね。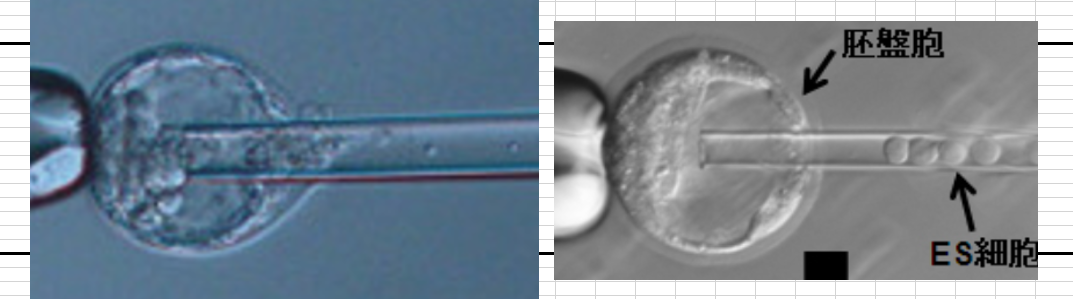
キメラと幹細胞は実際に存在しています。どうやって作られたかの可能性は以下の3つです。
①キメラと幹細胞は論文通りに作られたのに、若山さんはそれを既存ESによる捏造であったとして取り下げた。
②キメラと幹細胞は小保方核使用ntES実験で作られたものである。
③キメラと幹細胞は論文とは別の、又ntESでもない、何か違う手法で作られたものである。
2012年初頭から始まった二度目の実験で、F1は「僕のマウス」を渡したと若山さんは証言した。ところがその時のSTAP由来2Nキメラのジャームライントランスミッション結果であるカルスキメラ子1~9のDNAからAcr-CAG-GFPが検出された。
太田ESは使われていないということが証明されている以上、この「僕のマウス」を渡したという若山さんの証言も嘘であることが証明された。
彼は129/SvX1と岡部マウスとのF1の赤ちゃんを小保方さんに渡したということが立証されたのである。
このことによって、2011年11月の最初のF1キメラマウス背景を若山さんの実験ノートと証言によって桂報告書が(「129/Sv×B6GFP」 が正しい)と書いている(B6GFP)は岡部マウスであったからこそ、その後のテラトーマからもAcr-CAGが出たのだと知れる。
テラトーマの上からその時にできていたF1の幹細胞を注射したのは若山さんしかいないことも立証された。このことは手記206Pの(「若山先生は光る精子で実験をしていました」)という記述と矛盾しない。そもそも岡部マウスを維持しているのだから当然ですね。記者会見ではアクロシンが発見される経緯を自然に見せるために岡部マウスにはあたかも気づきもしないというポーズで、遠藤氏が言い出すまで知らんふりしていた。そもそも最初に疑うべきは太田ESではなくて岡部マウスでしょ。太田ESだと言いだして4種の細胞を取り寄せたのは若山さんです。
太田ESを若山さんに気づかせることなく小保方さんが渡すことは不可能であると証明された以上、太田ESはキメラ作成にも幹細胞作成にも使われていない。しかし、現実には桂報告とBCA報告はFES1=FLS3=129/GFPESとしている。太田ES即ちFES1は使われていない。これは何を意味しているのか。FLS3=129/GFPES=FES1ということである。
若山さんがFES1ラベルのチューブの中身をFLS3に入れ替えたということが立証されているのです。この件は早くから和モガさんが近親率から導いている結論でもありますね。
太田細胞が桂調査チームにもたらされた輸送経路こそ、今Ooboe さんのパートナー氏が神戸地方検察庁特別刑事部で検討していただいている申告書の嫌疑です。すべては山梨大若山研を経由している。(幹細胞のTCR再構成)小保方さんの酸浴細胞にはTCR再構成がありました。白線を入れてないルール違反だと大騒ぎしましたが、TCR再構成があることは石井さんも認めましたね。
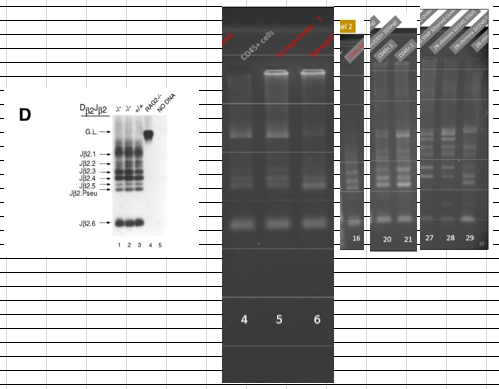
レーン4がCD45(白血球の分化抗原)陽性細胞、5と6がSorted Oct4+細胞、つまりその酸浴STAP細胞でOct4-GFPが蛍光しているものです。すべてのリアレンジバンドの出ていることが確認できる。多種のTCR再構成を起こしているT細胞をたくさん含んでいるポリクローナルな細胞集団ですから全バンドがでるんです。小保方さんのプライマーはD2-J2.6で挟んでいる。出るべきバンドは以下でしたね。
>>
01.D1-J1の123456-D2-J2の123456(*GL)
02.D1-J1の23456-D2-J2の123456(*GL)
03.D1-J1の3456-D2-J2の123456(*GL)
04.D1-J1の456-D2-J2の123456(*GL)
05.D1-J1の56-D2-J2の123456(*GL)
06.D1-J1の6-D2-J2の123456(*GL)
07.D1-J2の123456
08.D1-J2の23456
09.D1-J2の3456
10.D1-J2の456
11.D1-J2の56
12.D1-J2の6
13.D1-J1の123456-D2-J2の234567(*リアレンジバンド)
14.D1-J1の123456-D2-J2の3456(*リアレンジバンド)
15.D1-J1の123456-D2-J2の456(*リアレンジバンド)
16.D1-J1の123456-D2-J2の56(*リアレンジバンド)
17.D1-J1の123456-D2-J2の6(*リアレンジバンド)
細かい話は端折ります。01~06まではGLバンドです。小保方さんのプライマーで挟まれた一番長い断片です。ここに入るのはこの他にT細胞以外の細胞もあります。T細胞でない細胞はTCR再構成を起こさないので遺伝子はそのままですから小保方さんのプライマーで挟まれると何も切断されていないそのままの長さで出ます。因みにES細胞だけの細胞集団だとGLバンドだけになります。
07~12までは小保方さんのプライマーの部分がTCRの結果切り捨てられていますのでPCRにかかってきませんから、バンドはでません。
13~17がそれぞれ長さの違うリアレンジバンドです。バンドはGLも入れて全部で6本出るんです。
これを前提にして、では幹細胞のTCR再構成PCR結果はどうかというと、これは発表されていません。丹羽さんが主導して、幹細胞とキメラのPCR結果は論文から外させたんです。なぜか。幹細胞は実物の写真がありませんから先にキメラから示しましょう。
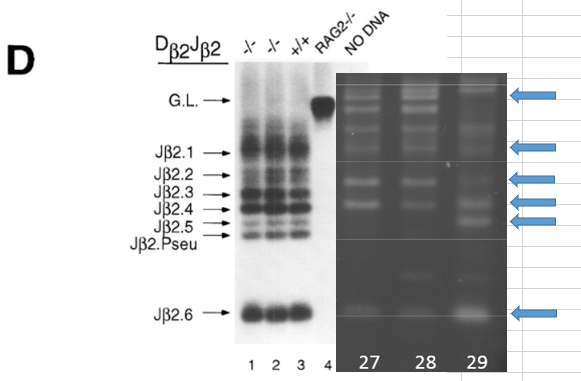
これはGel2のレーン27,28,29の2Nキメラの体細胞のTCR再構成結果とされているものです。27,28に関して上から5番目の矢印のバンドがありません。キメラの場合はポリクローナルな細胞が一つの組織の中に均等に分散するということは考えにくいんです。又、この実験自体がリシピエントの白血球を完全除去できているかどうかも分からないんです。で、議論を呼んでいるところですが、幹細胞の場合をこのバンドの欠失しているところから想像力を働かせることができます。
論文のプロトコル通りに幹細胞を作ったとしましょう。上に示したようにCD45+細胞と酸浴STAP細胞のTCRバンドは全バンド出ている。ポリクローナルな細胞集団だから全バンドが出るわけです。それをそのまま論文にあるES培地で誘導しても、細胞集団がポリクローナルな集団であることは変わりません。つまり作り立ての幹細胞であれば全バンド出るのが普通で、これが全く出ないとか、或は一部しか出なくても、検証している人たちはおかしいと思うか、目的のバンドは出ていないと判断しなければならない。全バンド出ない限りはTCR再構成があったとは言えない。
では、実際にはどういう証言になっているかというと、手記によればまず幹細胞のPCRはラボの仲間が担当した。そして、結果は8株のうちに2,3株あるようだと聞いたような書き方になっている。この時点ですでにおかしいですね。全ラインに全バンド出るのが当たり前です。これを小保方さんは当時気づかずにラボのミーティングで、自分のSTAP細胞の結果と合わせて、TCR再構成はあったと報告したようです。後にラボ仲間の実験にはコントロールが無かったので自分でもう一度やり直したら全ラインなかったと手記に書いている。この"後に"の時期がいつのことなのかははっきりとは書かれていない。それから全ラインなかったという意味はそれぞれの株で全バンドが出ていないから無いと言ったのか、全ての株で一切バンドがなかったのかのどちらなのかも書かれていない。しかし、前者であっても実験結果としては無いという判断が正しい。なぜなら検体はポリクローナルな細胞集団であるという前提になっているからです。
丹羽さんは実験が何らかの理由で正しく行われていないのではないかと考えて、間違いなく全バンドの出ているSTAP細胞の結果だけをつけさせて、幹細胞とキメラの結果は外させたんです。キメラは2,3種類程度の移植細胞が調査した組織の中でキメラ化していると考えれば説明できないことはないが、それもちゃんと学会で承認されている事実でもないし、何よりも特許につけられている2Nキメラの結果はもっと説明のできないものです。実験は4Nで行われた方が確実でもありますから、この時点では後にちゃんと調べればいいと考えて、キメラと幹細胞に関しては論文に載せないことにした。そして幹細胞のTCR再構成のPCR確認バンドが無くなった理由として、笹井さんは培養によるバイアス選択だと説明した。若山さんが嘘をついているなんてことは考えもしていませんから、他に考えようがなかったということです。
我々は若山さんが嘘をついていることを知っている。小保方さんの渡した酸浴細胞塊をそのままES培地で誘導したものでないことを知っている。
我々のntES論だと、作られてくるいくつかの小保方細胞核使用ntESは一個の細胞のクローン胚からES化したものですから、一つ一つのシャーレの中はモノクローナルな細胞集団です。
検体がモノクローナルな細胞集団である場合にPCRにかかってくるTCR再構成バンドは0本か、1本か、2本のいずれかです。こうなるのは小保方さんのプライマーには挟まれない断片があるのと、無論染色体が二本あるからですね。
8株の中の2、3株に再構成があるようだったというのは、1本とか2本なら分かりますね。0本もあるでしょう。でも、前提となっているのはポリクローナルな細胞集団です。1本や2本あってもあったことにはならない。それは何かの実験上の手違いでしょうと解釈されるべきですね。丹羽さんと笹井さんが心配して外させたのはそれが理由です。彼らは医師でもある。小保方さんや、ラボ仲間はそんなにTCRに詳しいわけもないんですね。後で若山さんに幹細胞と4Nキメラを作り直してもらって、自分たちが指導してもう一度確認すればいいことだと考えたんですね。とても正しい判断だったのではないでしょうか。若山さんが途中で逃げただけです。
以上
- 2019/08/31(土) 10:42:12|
- 一枚報告補記
-
-
| コメント:0
『一枚報告補記』(決定的証拠)以下の図はArticle Figure 4-aである。ここに若山さんの嘘を証明する写真が残されていた。
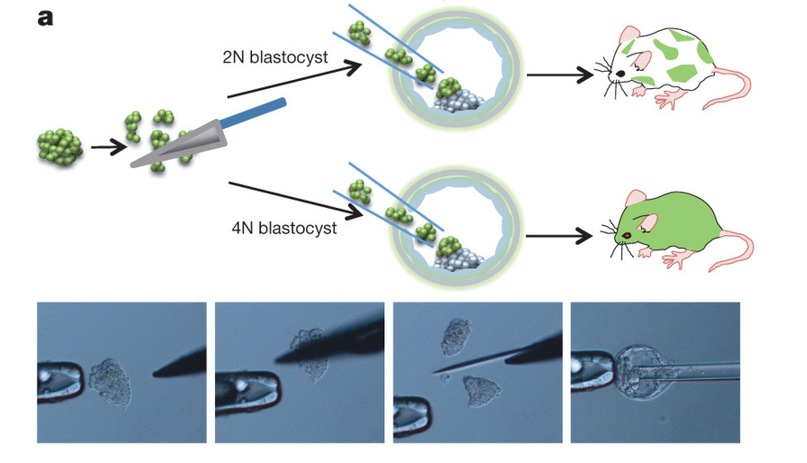
キメラ作成の概念図の下に、実際に若山さんが実験を行っている4枚の写真がある。左の端から、小保方さんが若山さんに渡したSTAP酸浴細胞塊、それをナイフカットしているところ、切り離された断片、そしてその断片をガラス管で吸い取ったものをキメラ作成のためのマウス胚盤胞にイジェクトしているところである。写真が小さいので気づかれにくかったが、以下が右端の写真だけを拡大したものである。
左がArticle Figure 4-aの下段に4つ並んでいる写真の右端を拡大したもので、右はNPO法人・発生工学研究会のホームページにあるES細胞の胚盤胞移植写真です。背景の2本の黒線は両方の胚盤胞の大きさを縮尺調整して同じになるように目安として私が入れた線です。マウス胚盤胞の大きさは直径ほぼ140マイクロメーターなので、黒線の間隔がその径となる。
左の若山さんの写真は拡大しているのでパイプの中やパイプの先にあるSTAP細胞の一つ一つが右側のES細胞と比較して如何に小さいかがよくわかる。
挿入用の管は胚盤胞を壊さないためにはできるだけ細い方が良いので、通常はガラス管を火であぶってES細胞の大きさに合わせて管をその都度作る。STAP細胞のナイフカット手法ではカット後の断片がES細胞の直径より大きいので太めの管を作っていることは左右を比較すると一目瞭然です。この太い管を挿入してそれでも胚盤胞を壊さないようにするには手技が必要となる。
笹井さんは2014/4/16の記者会見での記者の質問に対して、小保方さんが既存のES細胞を若山さんに渡して騙そうとしても大きさが違うからすぐわかると答えています。
左の写真の管の中の後ろの方にばらけたSTAP細胞が2,3個見える。右のES細胞と比較すると直径で半分以下です。直径が半分ですと面積では1/4です。実際そのように見えますね。でも体積は更に1/8です。左の写真のパイプの先に挿入した小さなSTAP細胞がたくさん見えますが、更に奥に大きな細胞が見えます。これがリシピエントのインナーセルマスなんです。その大きさは右側のES細胞とほぼ同じだと見えます。もともとES細胞というのはこのインナーセルマスを取り出して培養増殖させたものです。大きさは変わりませんが無限増殖します。
笹井さんはES細胞だったら若山さんは気づくといいましたが、笹井さんの記者会見の2か月後の2014/6/16に行われた自身の記者会見で、若山さんは記者の質問に答えて分からなかったと証言しています。やり取りの詳細は以下です。
>>
(全録)STAP細胞論文の共著者・若山照彦教授会見 質疑応答(2/4)
8:30/42:12~
(朝日新聞岡崎女性記者)
で、先生は実際に、その、STAP細胞からキメラマウスを造ったり、その、幹細胞を作成されたりして、実際に、その、細胞を見てるわけですけど、見た目、外観からすると、全く今までと違った細胞で、あの、他の類似した細胞というのは見たことなかったのでしょうか。
(若山さん)
マイクロマニピュレーターの上に細胞を乗せてしまって、そこからキメラとかを作るわけですが、その状態になってしまうと、あと、あの、今回に関しては、それまでずっと失敗続きだったということもあって、ええ、普通、キメラマウスを作るときは、細胞をバラバラにして、キメラを作るんですが、塊のまま入れてみようという、そういうアイデアで実験をしたということもあり、いつもと違う手法を取り入れた、その時に成功したんです。そのために、その時の細胞が以前と違っていたかどうかというのが分からないままなんです。
(朝日新聞岡崎女性記者)
では、その、もしかしたら、その、万が一ES細胞だったとした場合、それを、これが新しい細胞ですと言われて、ほんとに、見分けがつかなかったんですかね。
(若山さん)
そうですね。その時点では、あの、新しい手法でやってしまったので、見慣れた外観は全くないので、どの細胞だったかということは区別できなかったと思います。嘘ですね。それを証明しているのが上の比較写真です。誰でもわかりますね。笹井さんの言ったとおりです。当時専門家も気づかなかった。一つには論文をまだよく検討できてないということと、Article Figure 4-aの写真が小さくて、この管の中に大きさ比較できるものが写り込んでいるということに気づかなかったんですね。専門家は無論ES細胞の胚盤胞移植実験はよく知っています。でもSTAP細胞の大きさに関して直感的な認識がなかった。笹井さんは論文を書き直してやっている時によく見ていますから彼だけはES細胞なら分かるはずだと思ったんです。STAP細胞の大きさに関してはArticle Figure 1-gとExtended Data Figure 8-eにも図が掲載されているんですが、小保方さんは形態だけの比較で写真の縮尺を変えているから、これも気づかれにくいんですね。以下はそれぞれの写真の下部に表示されている1マイクロメーターのスケールの長さを合わせて比較したものです。
これも一目瞭然だと思います。笹井さんは小保方さんと机を並べてプロジェクターで図表の一つ一つを確認しながら論文をチェックしてあげている過程で大きさに関しての比較認識を持っていたわけです。ES細胞を浮遊培養して作ると見分けがつかないとか言葉だけであれこれと推測を述べていた専門家たちはこの大きさの問題には、本物を見たことが無く、直感を持っていないから気づかないんですね。
(推論考察の前提)若山さんは嘘をついています。そしてその嘘は最初のキメラ成功時の話にまで及んでいます。彼は小保方さんに対して当時ナイフ切り分けでできたと言った。そして事件化後には小保方さんにES細胞を渡されたと主張した。でもES細胞を渡されていたらパイプの引き延ばしはどうしたのでしょうかね。STAP細胞の挿入写真ではパイプの中に数十個は入ってます。同様にES細胞の塊をナイフカットして数十個の大きさにしたら胚盤胞には挿入できないほどのパイプの大きさになりますよ。つまりES細胞はそもそもトリプシンでばらして一個ずつ吸い取ってからでしか入らないんです。裏返すとテクニックこそ必要であれ、曲りなりにでも数十個の塊で入れられるということはどんなに個々のSTAP細胞が小さいかを知っていたということです。若山さんの嘘は明白ですね。
ここで最も大事なことは、小保方さんが既存のES細胞を若山さんに気づかれることなく渡すことはできないと証明されたことです。既存ES細胞を使ったのなら犯人はむしろ若山さん自身だということになる。ただ、若山さんがそんな単純な捏造をするはずは無いが、実際には何をしたからキメラができたのだろうかという方向に推論が行かないといけないのに、若山さんは既存ES細胞で捏造なんてするはずはないから、既存ES細胞を使ったのなら犯人は小保方さんだと堂々巡りする。若山さんに気づかれずに小保方さんが既存ES細胞を若山さんに渡すことはできないのだと、今証明された以上、既存ES細胞なら若山さんは犯人ではないというところに戻ってはいけない。既存ES細胞で捏造されたのではないというところに、まず、思考の固定軸を作らないといけないんです。桂報告書及びBCA報告の結論は間違っているのです。
(キメラは論文通りに作られたのか)思考の固定軸は既存ES細胞を渡されたのだという若山さんの主張は嘘だということです。ナイフ切り分けしたからできたと自分は思っていたが、実際には既存ESを小保方さんに騙されて渡されていたからできていたのだと主張していることは、今や嘘だと判明した。しかし、現実にキメラはできている。既存ESでないことは今証明されたのであるからには、キメラは論文通りに作られていたか、さもなければ別の手法で作られたのである。
この決定的な証拠の出る前には3つの可能性が検討されていた。
①小保方さんが若山さんに既存ES細胞を渡したからキメラができた。
②キメラは論文記載の通りにできている。
③キメラは小保方細胞核使用ntESで作られた若山さんの別の実験のものである。
①は完全否定されて、桂報告書とBCA報告の結論は間違いであると証明された。②と③のどちらが正しいのかが残された問題です。
キメラは論文通りにちゃんとナイフ切り分けでできていたのだけれども、自分自身が特許の件に不満があったとか、iPS細胞やミューズ細胞との予算獲得競争者や裏についている製薬会社等の圧力や脅迫に負けた等の、私にはわかりませんが、何らかの理由で若山さんが論文を取り下げようとして、キメラは既存ES細胞を渡されたからできたのだと嘘をついたという推論と、そうではなくて若山さんは論文通りの方法ではキメラを作っていないのに、笹井さんの圧倒的な信用力で間違った論文が通りそうになったので、全てを小保方さんの所為にして、論文を取り下げようとしたのだという私の説です。
私の説を概括しておくと、若山さんはスタンダードなプロトコルではキメラができないということを確認した後、小保方核使用ntESからキメラを作ることによって、小保方細胞の性質を見極めようとする別の実験を計画していた。そして米国に帰って、自分の研究を続けようとしている小保方さんを、人事秘の解ける翌年初頭に山梨大の助手で採用するという条件提示ができるまでの間、一時的に日本に引き留めておくために、自分のntES化実験で作ったキメラをなぜできたかを語らずに、むしろナイフ切り分けでできたと嘘をついたのだという説です。
以上
- 2019/08/27(火) 14:55:28|
- 一枚報告補記
-
-
| コメント:0
(2) 疑念の検討A. コントロールES細胞a.AC129実験と多能性遺伝子解析実験の区別小保方さんはF1の受精卵ESは若山さんにもらった「僕のマウス」ESしか持っていません。それ以外に持っているのは学生のGOFのntESだけです。そして桂報告書によって太田ESを持っていたのではないかと疑われているわけです。しかし、ここでは太田ESは使われていないから関係ありません。
AC129というのは若山さんが129/Svの赤ちゃんマウスを小保方さんに渡して、STAP細胞を作ってもらって、それを培地誘導で幹細胞化させたと言ってるものです。129/Svを渡したのにAc129-1を遺伝子解析したら129B6F1CAG-GFP(ホモ)でしたという調査報告です。しかも、遺伝子欠失と重複の特徴からして、129B6F1CAG-GFP(ホモ)マウスから若山さんが作ったESの内のライン1だったということです。
それに対してSTAP ChIP lysateは、そもそも、今は雌雄表記の問題はわきに置いておいて、B6 x 129/SvのF1マウスだと小保方さんがデータベースに書いているのですから、129/Svの背景が出たらむしろおかしいわけです。GRASに提出して調べてもらっているのは多能性遺伝子の発現状況です。129/Svの実験とは基本無関係のはずなんです。
どうして調査チームはAC129の実験サンプルと多能性細胞遺伝子解析実験サンプルを混同したのか。それは竹市さんが言ったように最初にちゃんと調査対象のサンプルの出所と由来を調査しなかったからです。証拠の信憑性も含めた調査対象サンプルの証拠能力調査という、警察の捜査なら常識である手順を踏んでないからなんです。
サンプル調査は2014年の3月10日に若山さんが論文撤回を著者たちに申し入れたその日に、調査対象被疑者であるはずの本人が理研の許可もなく勝手に第三者と称する放医研の知人等にサンプルを送った。恐らく守秘を配慮して若山さんがMTAを結ばずサンプルを山梨大に持ち出さすことを積極的に依頼した手前、事件化して後は山梨大に依頼してサンプルをすべて差し押さえなければならない理研がMTAが無いことによって依頼できなかったという弱みをにぎられていたのである。さもなければ若山さんは試料を差し押さえられて勝手に解析に出すことはできなかったはずである。
その時に放医研に出されたのは以下です。
①FLS 8株
②FLS-T 2株
③AC129 2株
④GLS 2株
⑤コントロールES 5株
全て山梨大で彼が保有していた試料で、従ってAC129も若山さんの持ち出していた試料を送っている。この結果が6月16日の記者会見で発表されてGFPの挿入位置が18番染色体にホモに入っていて、コントロールESと同じだと分かった。調査は若山さんの主導で始まっていて、証拠能力の検証が先だと言った竹市さんと野依さんの意見は通らなかった。
AC129からF1背景が出たからAC129とF1が関係づけられて比較分析対象となったが、今残されているAC129が当時129/Svで行われたと言われている実験とどういう関係なのかということは調査されていない。ただ若山さんの証言を証拠もなく妄信しているだけである。ここに大きな疑念があるが、今は、細胞比較をリヴューすることから始めよう。
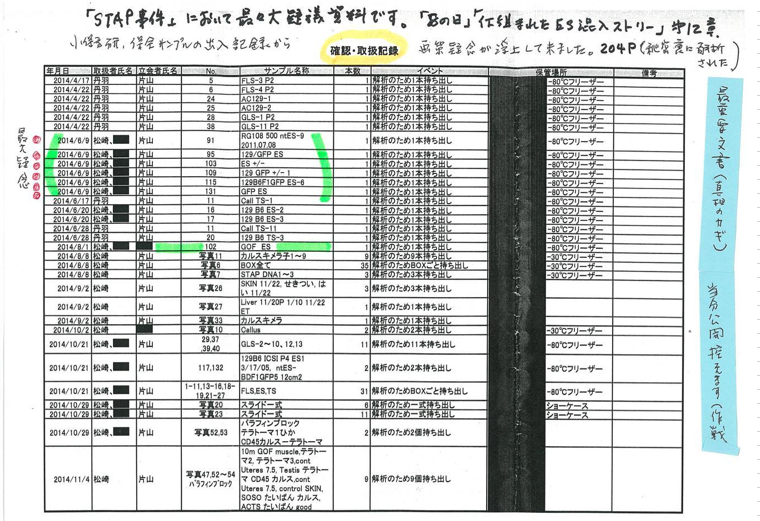
b.129マウスコロニーの不均一まずAC129が全解析されて、次に小保方研フリーザーにあった129B6F1 ES6が怪しまれたがために全解析を受けたと思われる。というのも仮に放医研から回ってきた129B6F1-1,2,3,4,6 ESの中の6だったら、これをまず全解析する理由はありません。佐藤貴彦氏の『STAP細胞 事件の真相』の中で、取り寄せた公開資料により、6月2日に以下の細胞が分析されていると書いています。又後のOoboeさんのパートナーさんの取り寄せ試料から管理室の持ち出し日も分かっています。
>>
①FLS-3(木星リスト5番 4/17 丹羽氏持ち出し)
②FLS-4(木星リスト6番 4/22 丹羽氏持ち出し)
③GLS-1(木星リスト28番 4/22 丹羽氏持ち出し)
④GLS-11(木星リスト38番 4/22 丹羽氏持ち出し)
⑤AC129-1(木星リスト24番 4/22 丹羽氏持ち出し)
⑥AC129-2(木星リスト25番 4/22 丹羽氏持ち出し)
①②はこのときに15番にGFPが入っていたとされたが放医研も理研も間違えた。GFPにアクロシンのプロモーターがついていたことからアクロシンの遺伝子は15番なので15番だと判断したが、これは岡部マウスのAcr-GFPの前につけられているアクロシンプロモーターで、実際には3番に入っていたことが後に分かったというお話になっているのはよく知られている。物事が順番に判明して行ったかの如くのお話になっている。ここで登場するヒーローもよく知られている人です。後程触れましょう。
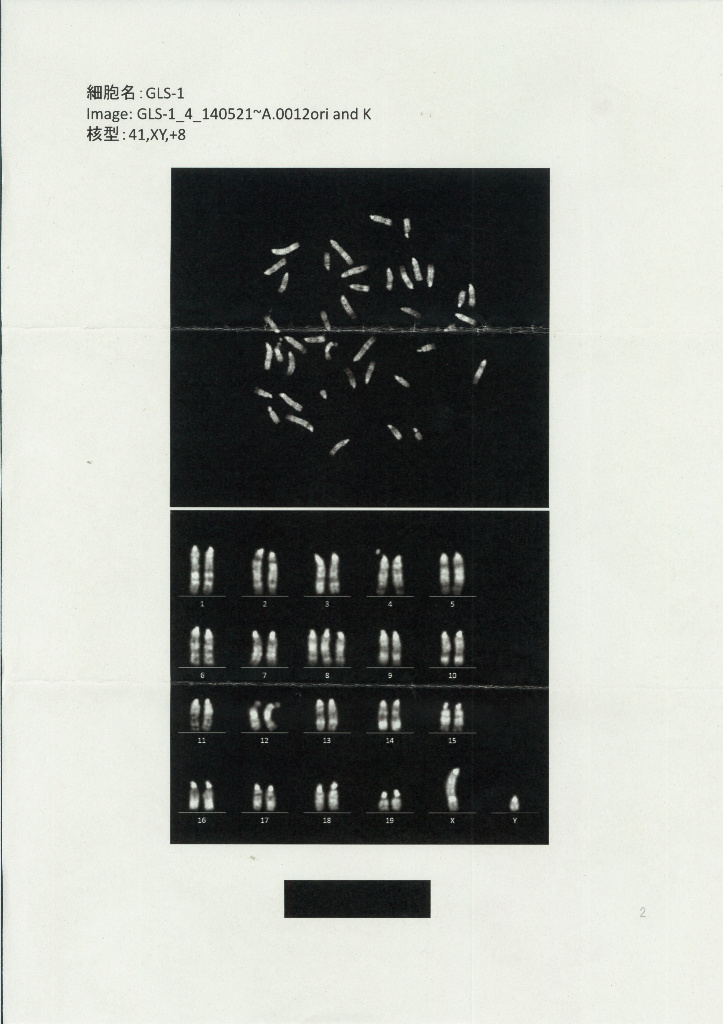
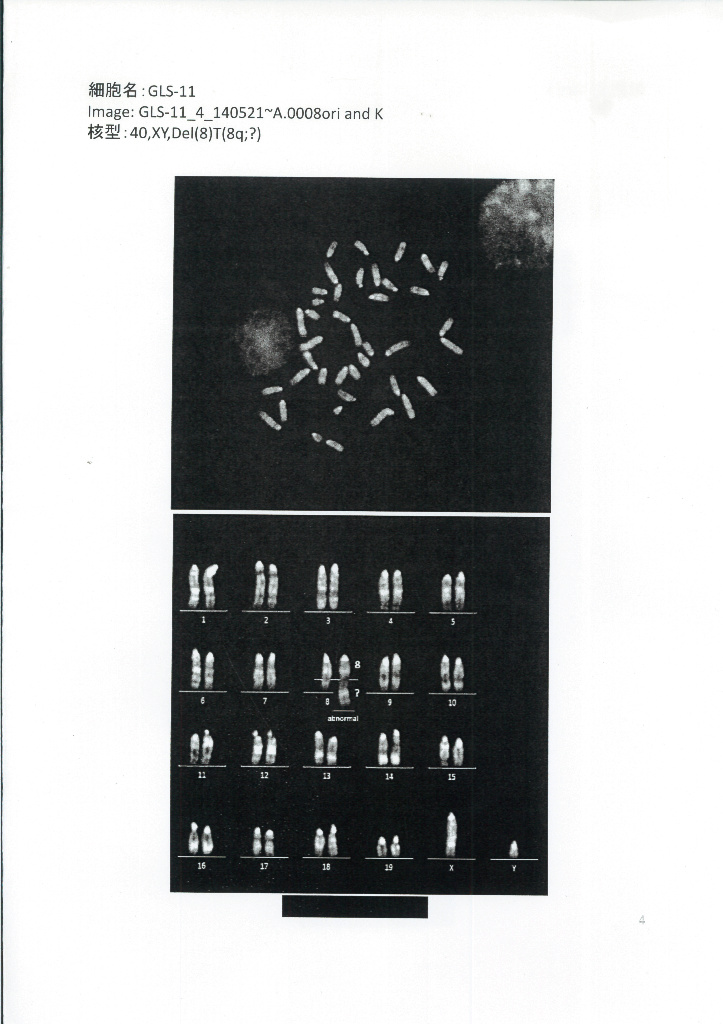
因みに③④については丹羽さんが核型解析に出したんです。DORAさんの昔のブログにある。保存のためにこれもここに貼り付けて置きましょう。



佐藤氏によるとこの時にはすでに以下のサンプルも解析されていたという。
>>
⑦TSA ntES-9(木星リスト91番 6/9 松崎氏持ち出し)
⑧ES129 GFP+/-(木星リスト109番 6/9 松崎氏持ち出し)
⑨GFP ES(木星リスト131番 6/9 松崎氏持ち出し)
⑩129/GFP ES(木星リスト95番 6/9 松崎氏持ち出し)
⑪ES+/-(木星リスト109番 6/9 松崎氏持ち出し)
⑫129B6F1GFP ES6(木星リスト115番 6/9 松崎氏持ち出し)
⑫が全解析されたんですね。AC129は既に丹羽さんによって4/22に持ち出されている。最初丹羽さんは細胞の解析にも参加していた。しかし、後に外されて再現実験の担当になります。6/28に持ち出したのが最後で再現実験に移り、後を受けたのが松崎氏です。
因みに本日(2019/8/19)、Ooboe さんが学さんブログに以下のように書き込まれている。
>>
・・・6月2日にはStap関連と129B6F1ES6、129/GFP ESなどが解析終了してます。が記録にありません。この事実は重いのです。
竹市所長のしぶしぶ了承は6月5日です。竹市所長Goの前に持ち出しているのです・・・
2019/8/18(日) 午後 6:12[ Ooboe ]返信するこの件の経緯に関しては佐藤氏の『STAP細胞 事件の真相』(2016/12/14初版発行)の80P前後に既に書かれていることですが、Ooboeさんのパートナーさんは後にサンプルの調査持ち出し記録を入手されたのでその情報も付加されている。どちらも6月10日付の竹市センター長の報告書を根拠にされているが、
その内容が開示されていないので、サンプルが持ち出し記録にある6/9ではなくて6/2以前だということの確証がない。無論両方で主張されているので、その竹市報告に書かれているのだろうとは思いますが、早く開示されて欲しいものです。
参考のためにOoboeさんのパートナーさんが取り寄せた調査用のサンプル持ち出し記録を保存のために貼り付けて置きましょう。二列目が持ち出し者の欄です。
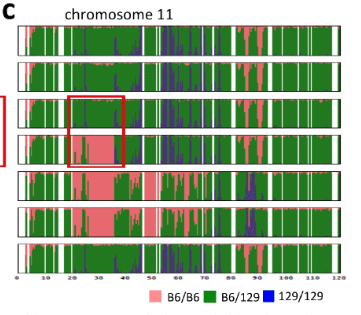
しかしこの時に全解析を受けた⑫はAC129の6番染色体にあった遺伝子異常と一致しなかった。そのため、ES1~5の6番染色体だけのシーケンスが行われたとみられる。このシーケンス結果が添付されてない理由はそれぞれがすべて微妙に違っていて、それを見せると一般の人たちから違うじゃないかと言われるからです。Extended Data Figure 2-cを再掲します。3度目の検討になります。
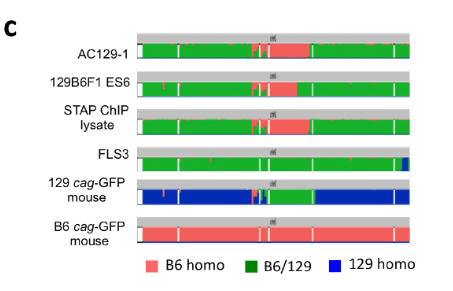
6番染色体です。B6マウスはすべてピンクです。129マウス中央にグリーン部分がある。ここはピンクとブルーが半々の場所です。ピンクとグリーンの掛け合わせからすべてグリーンの部分はできません。逆に言うとF1側でグリーンになっている場所は129側ではブルーだということです。
ここで分かることは上図に挙げられている129 cag-GFP mouseのSNPs分布はいろいろある129の中の一つに過ぎないということです。129のマウスコロニーは均一ではないのです。均一でないマウス同士の掛け合わせが行われて「僕のマウス」ESが作られている。均一だったら二種類のSNPs分布のESしかできない。それが1~6まですべてわずかに違っているんです。それは129マウス自体に何種類か混じっていてESが作られるとき複数のペアから採卵されている可能性もあるということです。桂報告書はこのことを以下のように説明している。9Pです。
>>
4)他の細胞株における遺伝的不均一性
この遺伝的不均一性は、129 CAG-GFP マウスに由来する他の細胞株にも反映していた。 若山氏により 129B6F1CAG-GFP マウスの独立した胚より複数の受精卵 ES 細胞株が樹立 されているが(129B6 F1ES1~6、2012 年 5 月作製)、いずれも第 6 染色体中程に B6 ホモ 領域を有していた。しかし、この B6 ホモ領域と 129/B6 ヘテロ領域の境界は 129B6 F1ES1 ~6 の間で異なっていた。このばらつきは、129 CAG-GFP マウスの配偶子が形成される 際、減数分裂の過程で、B6 と 129 の染色体の組換えによって生じた可能性が高いと考え られる。ただし、細胞株樹立時の体細胞分裂における染色体組換えがこの多様性に寄与 した可能性もある。ES1~6ですべて微妙に異なっていたことを認めています。素人に説明するのは大変だと見て意図的にシーケンス結果を並べなかったんです。
そして見せないままにその原因を減数分裂時と体細胞分裂時の染色体組み換えに求めている。しかし、そんなに都合よく微妙な場所だけで組み換えが起こるということは考えられない。
そもそも大きなヘテロ領域があると減数分裂で基本的に二種類できると先に説明したことを忘れている。SNPs分布でAC129 typeとES6 typeに分けましたよね。本当は2種類どころではなかったということです。
マウスコロニー自体に微妙な不均一が入っていて複数種のペアから採卵されているとこうなりますね。若山さんの維持している129CAGホモは長く維持しているからそれなりに近交系マウスになっていて維持メイティングはインクロスになっているはずだということを上述しています。途中で又B6が飛び込んでいるんですね。事故コンタミですね。気づいて慌てて取り出してもすでに交接してたということでしょう。無論、自家繁殖ですからこういうことはあるんで、実験の目的に支障なければ問題ないんです。若山さんの実験室では困るような事態では無かったということです。小保方さんの実験に使って、捏造事件になったから問題になっただけで、逆にマウスのコンタミが無かったら識別できなかったでしょうね。そもそも製薬の実験なんかに使われるマウスでこんなことがあったら大問題で、だから自家繁殖させてはいけない契約になってるわけです。若山さんはただ自分のところで使ってるだけです。まあ、予算節約ですね。まさか予算請求して懐に入れると飛んでも無いことになりますからそれはやってないでしょう。予実対比されますからね。経理部門がチェックしている。経理部門が何かしてるとまた別問題で、今度は会計監査が問題になる。そして公的部門には国税は入らないですからねえ。はは。
強引な結論に導こうとしてデータ開示しないからこういう訳の分からない説明になるわけです。cのリジェンドに以下のように書いていますが、書いている本人がクッソーと思いながら書いてるんじゃないですか。
>>
Note that the 129/B6-heterozygous SNP region in the 129 cag-GFP mouse is longer than that of AC129-1.
[ カツラ報告書 ]氏が指摘された場所もそれが原因です。
c.MTA無しの山梨大への移籍若山さんは2013年3月に理研内ラボを引き払うに際してMTAを締結しなかった。これは既に笹井さんが参加していてヴァカンティと特許の件に関しても話がついていて、米国側から守秘に関する依頼があったものと思われる。MTAを締結すると山梨大側にその情報が漏れてしまうということがあって、本来なら締結が必要であるが、論文発表まで待って事後締結すると打ち合わせられていたと推定される。手記に事務方から若山さんがMTAをなかなか出さないので訴えるという話を小保方さんが聞かされているのは、無論、イレギュラー処理をしていることを小保方さんに隠そうとしているので、若山さんとは打ち合わせ済のことである。こういう事務処理はマニュアル化されているので忘れるなどということはない。意図的に行われているのである。
しかし、このときにイレギュラー処理したがために、事件化後、理研は山梨大への若山ラボの冷凍サンプル保全命令依頼をできなかった。MTAも無いので山梨大はこのことを全く知らないからである。その結果、若山さんは理研の調査が始まっているというのに被疑者であるにも関わらず、自らが潔白であることが当然であるかのことくにサンプルを勝手に分析に出すという振る舞いができたのである。
2014/4/1に事後提出されたMTAの添付持ち出し資料にはコントロールESはES1しか持ってないことになっていて、3/10に放医研に出したコントロールESが5株であることは既述しています。調査体制の調整が何ら取られていないということも申し上げた。
この事後MTAに関して開示請求を行ったのが木星さんです。その後、このリストを使った持ち出し記録を開示請求したのはOoboeさんのパートナー氏です。
小保方さんが2012年8月にGRAS提出してLetter Figure 4-bで使ったESは以下の木星リストの131番ではないかと疑義されます。これはおそらく「僕のマウス」ESの1と6ではないか。

若山さん由来のGFP ESと言ったらコントロールの「僕のマウス」ESしかありません。小保方さんの略し方で129B6F1は129で、129ESと書いたらFLSのことでESはES-likeだと分かっている。
こういう書き方は無論FLSという名称が決まる前の状態でのサンプルです。STAP細胞が最初はスフィア→カルス→アニマルカルス→STAP細胞と変遷していくのと同じです。FLSという言葉が与えられる前はES様細胞と考えられているわけですからES-likeで、小保方さんはまだF1のコントロールESが作られていない時期ですから129ESと書いたら129B6F1のES-likeと自分で分かるからそう書いている。コントロールESを貰った後もGFP-ESと書いたらそれと分かる。学生から貰ったGOFのntESに関してはGOF-ES1と書かれていて、調査時に(■■由来→小保方)と答えていているのですぐに識別できているわけです。
でも、小保方さんは調査時には何が起きたのか全く分からない状態ですから、先生のラボに迷惑を掛けることもできない中で、口ごもりながらおどおどと答えることになる。調査側の考慮がまるで無い。周りが保身で汲々としている状態のようですね。こういう状態だと生贄の山羊になり易い。
GFP ESと書いてしかも若山さん由来だと、これは本当のES細胞です。これが1番と6番だったものを何度も解凍凍結を繰り返してラベルも書き換えられたものではないでしょうか。2本ある。旧DORAブログにある写真です。

これでは若山さんには気づけませんから若山さんは6は無いと思って、117番を忍び込ませたのではないか。松崎氏は131番を持ち出して調べているにも関わらず結果報告をしていない。
謎めいたところですが事実は以下です。
>>
理研小保方フリーザーにあるF1ESは以下です。
129 B6 ES-2
129 B6 ES-3
129 B6 ES-4
129 B6 ES-5
129B6F1GFP ES-6
GFP ES
若山さんが放医研に出したと言ってるF1ESは以下です。
129B6F1-1 ES
129B6F1-2 ES
129B6F1-3 ES
129B6F1-4 ES
129B6F1-6 ES
若山さんの事後MTA締結添付持ち出しリストにあるF1ESは以下です。
ES細胞 129B6F1 GFP-1
d.引っ越し時のサンプル整理上述した131番の下の132番はntESのBDF1ですから若山研で本来行われている実験のサンプルが紛れこんでいる。小保方さんは当然「不明」と答えています。
因みに若山さんの奥さんが警察の調べに自分のものと言ったと小保方さんが聞いたという日記の話はその132番と以下の117番です。それぞれ1本ずつの計2本です。

DORAさんの旧ブログに写真がある。GFP ESの2本も見えます。彼女の字です。若山さんのだとすぐわかったのだから自分の字なんです。このSの字は特徴がありました。P40 FLS-1~8のSです。彼女はちゃんとFLSの40継代120日間を実施しているんです。
小保方晴子日記は以下です。
>>
2016年
2月16日(火)
・・・私が新しく聞いた話もある。窃盗の疑いがあるというES細胞についての説明を受けた。報道された元留学生が作製したES細胞に加え、「ヒッポさんのもの2本」と写真を見せられた。その写真は理研の調査委員会の調査期間中、「持ち主不明のES細胞」と呼ばれていた細胞チューブだった。私は見知らぬES細胞が自分のサンプルボックスに入っていたことを知らされ、薄気味悪く思っていた。結局、「持ち主不明」と呼ばれたまま、調査は終了した。ともに公の場であるにもかかわらず、調査委員会で知り得なかった新事実を、事情聴取の場で聞かされた衝撃は大きかった。思わず刑事さんの話を遮って「ご本人の証言ですか?」と聞いてしまった。「そうです」と返事された。
ヒッポさんのだったんだ。盗まれたって言ったんだ。
ビニールとかプラステチックとか、そういう消化できないものを飲み込んでしまったみたいに、まさに消化できない気持ち悪さが残った。松崎氏は持ちだして調べています。報告がない。盗まれたと奥さん自身が警察の尋問で答えたのかどうかは日記の記載だけではわかりませせんが、石川氏がフライデーで盗まれたのだと書いたのは間違いありません。彼は山梨に訪ねて行って、若山さんと会ったが若山さんは体調が思わしくないので、奥さんと大日向氏が石川氏に説明したのでした。
因みに小保方さんの筆跡は以下です。日経サイエンスにこの写真が流出したのは調査に関与していた松崎氏のラボからです。P40のFLSが1~8までそろっているのに皆でスルーして、増殖率表の捏造だと騒いだんです。何なんでしょうか、この人たちって。
FLSのSの字を上のGFP ESのSの字と見比べると同じ筆跡と分かる。
40パッセージが終了したのは120日後の5月末頃ですから、その時にはすでにFLSという言葉が作られている。

 e.若山さんによる捏造
e.若山さんによる捏造このAC129とFLS-Tシリーズに関する捏造と他の捏造とされているもの(我々の説では岡部マウスとのF1マウス等を使った小保方酸浴細胞核使用の正規のntES実験)とは違うものです。
最初のキメラ成功から翌年初頭の確認実験や続く胎盤実験とCTSの作製までは、そもそも小保方酸浴細胞核使用ntESの性質解明実験で、実験自体は何も捏造実験ではなく、F1に関して使われているB6は最初からアクロシン入りの岡部マウスであっただけの話です。
太田ESや学生が渡したGOFのntESを小保方さんが捏造させるために若山さんに渡したのだという話は、若山さんが最初リクルート上の理由があって、小保方さんを引き留めるために、本当のことを言わずに、スタンダードな手法でキメラができたかのように小保方さんに話していて、ヴァカンティまで本気にさせてしまったので、いざというときには自分のESコンタミで言い訳しようとしてヘテロの話を造って、その伏線も小保方さんに吹き込んでいたものです。最初から小保方さんの所為にしようとして作った話ではなく、自分のミスだったと言い訳するための作り話に過ぎなかったものだ。無論実験自体は何も捏造ではなく、本当のことを小保方さんに言ってないがために論文が嘘になっていただけだ。若山さんは通らないと分かっていて、取り敢えず論文を書かせれば小保方さんはヴァカンティから自由になれると思っていた。それが本当のことを言わないままに小保方さんに論文を書かせていた理由です。小保方さんを理研が引き抜いた時から若山さんが事実を正直に言わない限り、いざというときのための言い訳として小保方さんの意図的コンタミというストーリーに変更せざるを得なくなった。道は3つに分かれていた。
①本当のことを言う(自分のプロモーションの話も含めて今更いえなくなってしまっていた。)
②論文のリジェクトを願う(専門家として通るはずはないと予測するのはそれほどおかしくない。そもそも三誌リジェクトされている。)
③小保方さんの所為にして逃げる(笹井さんの圧倒的な信用力で通ってしまいそうになり、現に最終的に通った。)
AC129の実験はまだ小保方さんが若山研に所属していて笹井さんが入る前の実験です。細胞の培養開始日は2012/8/13です。小保方さんは129/Svの赤ちゃんマウスを2012/8/5前後に若山さんから受け取ってSTAP細胞を作った。この実験がとても不可解なものでした。
f.混乱期の状況それを考える前に、この頃の状況を確認しておくと、小保方さんがプイと米国に帰ってしまったのは手記によれば、2012/11/15の数日前ですから2012/11/10前後ですが、小保方さんはこのAC129の実験が開始された2012/8/5頃の丁度3か月後にヴァカンティの許に帰ってしまった。つまり山梨にはいかないと返事したことになる。この返事の仕方は手記を読む限りでは、とても失礼なマナーです。ちゃんと筋を通すことなく、寺下さんにアメリカでやりたいと内心を打ち明けて渡米した。若山さんの耳に入ることを承知で打ち明けたでしょう。無論、渡米した以上、ヴァカンティや小島、大和に事前に相談している。そのことも手記に書いてある。しかし、推測では米国から小島を通して共同研究解除の話をしていると思われる。その後すぐに西川さんから理研への勧誘の話が入って、結局ちゃんと本人が挨拶してない形のままではなかったかと疑われる。直後に理研の話が入ったので気づかれにくいが、理研が誘わなかったら、小保方さんは若山さんとヴァカンティの間でヴァカンティを選んだという形になる。結構、この形は失礼でしょう。よほどちゃんと大和や小島と一緒に訪問して、自分の気持ちを説明し、お礼を言ってヴァカンティのところに戻るということをしないと、若山さんのプライドを痛く傷つけたと思います。日本とは学制が違うとはいえ、ヴァカンティは一介の麻酔医で博士号も取得していない人だ。方や、若山さんは世界初のマウスのクローン作製とntESの作製に成功している世界的な学者だ。記者会見で一度ヴァカンティと呼び捨てにして、後にヴァカンティ先生と言い直しています。特にリクルート絡みで対抗していましたからあまりしっくりした仲ではないでしょう。まあ、大人ですから態度にはださないでしょうが、結構ムッと来るような仕儀だと第三者が読んでいても感じるところですね。「残念だけれども山梨大の助教よりも理研のユニットリーダーを選ぶのは当然の選択です。面接頑張ってください」と励まされた若山さんの言葉を書き留めているんだけれども、理研が誘わなかったら、一介の麻酔医であるヴァカンティ研で一介のポスドクであることを選ぼうとしたのがあなたの当然の選択なのだろうかという皮肉を、小保方さんは感じたでしょう。若山研の客員に決まった経緯の中で小保方さんは若山さんに論文が出るまではヴァカンティ研に所属してくれと言われたと説明しています。手記にちゃんと書いてある。それはヴァカンティに言われたとおりのことなんでしょうけど、若山さんは論文が出たら彼女のリクルートは可能だとなんとなくは思っていたでしょう。そして、キメラは出来なかった。帰ろうとする彼女を翌年の山梨大助手の条件提示できるまでの時間稼ぎとして、別途考えていた酸浴細胞核を使ったntES化実験でできたキメラを説明なしでできたと一時的嘘をついた。そして翌年に助手で誘ったけれども彼女は即答しなかった。これでどんどんこじれて行ったんです。人はそれぞれ思いが違います。彼女は女性ですからあまり地位に拘りません。男性みたいな権力欲が薄い。米国の空気は自由なんで結局彼女がヴァカンティのもとに戻りたくなったのはそれが一番の原因だと思います。それに対して若山さんは米国の過酷な競争社会を知ってるし、父性から自分のところで落ち着いて成長させてやりたいという気持ちもあるんでしょう。こういうのってドモナランですなあ。世の中のことは誰でも自分の思い通りにはならない。それで運命という言葉もあるんでしょうや。
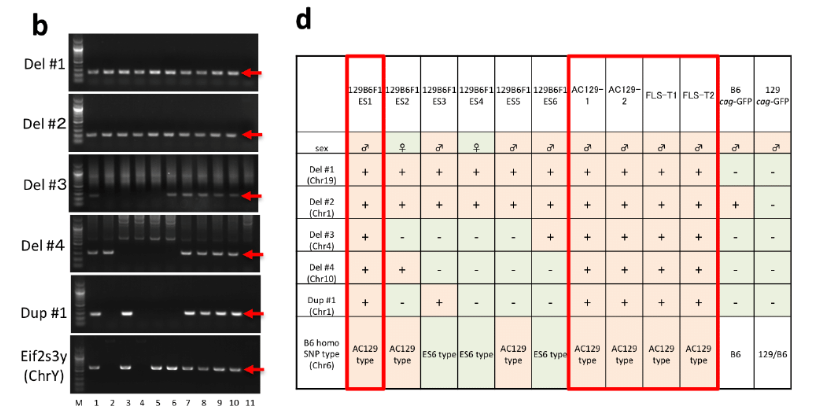
B. 欠失と重複a.全解析まずBCA報告のExtended Data Figure 2-c,dを再掲します。
bとdのリジェンドは以下です。
>>
b, PCR detection of chromosomal anomalies and Y chromosome in cag-GFP STAP stem-cell lines and parental mouse strains. Lanes 1–6: control ES cells, 129B6F1 ES1–6; 7: STAP stem cell AC129-1; 8: STAP stem cell AC129-2; 9: STAP stem cell FLS-T1; 10: STAP stem cell FLS-T2; and 11: GOF-ES. Deletions 1–4 and duplication 1 are located on Chr19: 32,857093–32866,121, Chr1:140,698,249–140,702,693, Chr4:123,747,239–123,763,596, Chr10:43,265,147–43,267,270
d, Table summarizing the chromosomal anomalies and differential types of Chr6 B6-homozogous SNP clusters in the cag-GFP cell lines and parental mice. Control ES cell 129B6F1 ES1 shares all the characteristic features with the four cag-GFP STAP stem-cell lines.この中でWGSに掛けられた最初の細胞サンプルはAC129-1です。桂報告書の8P。
>>
小保方研フリーザーに保管されてい た STAP 幹細胞 AC129-1 について、SNPs マーカーの TaqMan PCR 法による解析を行い、さ らに NGS により全ゲノム DNA 配列を解析した。同じく STAP 幹細胞 FLS の対照として CAG-GFP マウスから作製された受精卵 ES 細胞(129B6 F1ES)の解析も同時に行った。まず最初にAC129が全解析されて、4個の遺伝子欠失と1個の重複が見つかった。BCA報告に貼付されているWGSに掛けられた細胞サンプルのリストは以下です。
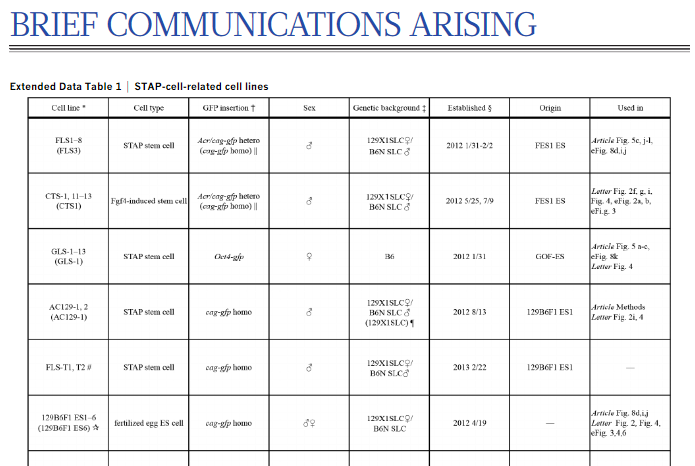
以下が関係したリジェンドです。
>>
*The line subjected to WGS is indicated in parentheses in cases in which several sublines were established for one cell type. Other sublines were confirmed by PCR and sequencing.>>
☆The genome of 129B6F1 ES6 was sequenced, but further analysis showed that 129B6F1 ES1 rather than 129B6F1 ES6 shares all genomic anomalies found in AC129-1.control ES 129B6F1 ES1はWGSには掛けられていません。掛けられているのは129B6F1 ES6で、他はOther sublines were confirmed by PCR and sequencing.とあるとおりです。でも、その結果は図示されていない。理由は既に述べた通り、全部違ってるからです。違うなら違う理由を説明すればいいだけですが、厳密にはわからなかったので、図は世間には見せないことにしてしまった。
それをごまかす為にb図の並びは4欠失と1重複、そして性別なのに、d図では性別を先にし、4欠失と1重複、そしてES1型とES6型の二種類のSNPsパターンに分類されるとした。図をつけると全部違うからつけなかったということです。ES1型をAC129型としているのは最初にAC129を全解析して、次にES6を全解析したが、遺伝子異常もSNPs分布も違っていたので、更にその後にES1~5の6番染色体だけをシーケンスしたためです。全解析されているのはAC129とES6だけです。AC129には4つの欠失と1つの重複があって他の染色体にはない。ES6には3つの欠失があって他の染色体にはない。ではES1~5の遺伝子異常はどうかというと全部は調べられてないということです。AC129の全解析で見つけられた以下の異常個所だけをPCRで確認したんです。
>>
①Del#1(Chr19)
②Del#2(Chr1)
③Del#3(Chr4)
④Del#4(Chr10)
⑤Dup#1(Chr1)
他の場所に有るか無いかはこの方法では分からない。ES1はPCRでAC129にあった遺伝子異常だけ調べて他は調べてないから同じだという証明は不完全ですが、結論は以下となっているのです。
>>
STAP 幹細胞 AC129-1、AC129-2、並びに、STAP 幹細胞 FLS-T1、FLS-T2 は、129B6 F1ES1 に由来すると結論づけた。 正しくは、現在のサンプルの中身はAC129-2、STAP 幹細胞 FLS-T1、FLS-T2 及び129B6 F1ES1 はSTAP 幹細胞 AC129-1の遺伝子異常と6番染色体上のB6と129のそれぞれの特徴的SNPs分布を
共有しているということです。
そもそも何がどう使われたかなんて、AC129-1の中身をAC129-2、STAP 幹細胞 FLS-T1、FLS-T2 及び129B6 F1ES1に入れてもそうなります。先にサンプルの証拠能力を調査検証してないから何事も言えません。全サンプルMTAもなく無断で若山さんは山梨に持ち出してるんですからどうにでもできる。
b.細胞の同定さて、まあ、それでもAC129-1の遺伝子異常は以下の4つのサンプルと共有されていたことは事実なんでしょう。
①129B6F1 ES1
②AC129-2
③FLS T1
④FLS T2
①のマウス背景は実物があって「僕のマウス」です。調べた結果分かっている。若山さんが一人で作って若山さんが持っていて、かつ小保方さんに渡したものもある。ただし、事件化後に残されていたサンプルの保管状況では小保方さんはES1をもって無くて、それは山梨で若山さんが持っていた。
②のマウス背景はAC129のCAGホモだと若山さんが証言しているし、白毛なので小保方さんも赤ちゃんマウスを渡されたときには1のアグーチとは違うと分かります。でも出来上がったものは「僕のマウス」だと調べがついてますから、若山さんが小保方さんの捏造と見せるために自分でマッチポンプの捏造をしたか、小保方さんが白毛のマウスを渡されながら、愚かにもひょっとしてキメラを作られたらすぐばれてしまうのに「僕のマウス」ESを渡したか、キメラは作らないと知らされていたので違う背景のESを渡したのどちらかです。
論文にはこの129/Svでキメラが作られたと書かれています。小保方さんが犯人だとこれは嘘になります。それと論文ではこの129/Svは129/Sv carrying Rosa26-gfp と書かれていて、これも小保方さんの嘘ということになってしまうんですけど、CAGホモと聞かされているか、もしくは何も知らされていないのにcarrying Rosa26-gfp と書くと、若山さんに嘘を指摘されるからそんなこと書かないでしょう。若山さんかもしくはラボの誰か、特に奥さんがそういったということで、この段階ではすでに小保方さんは嵌められようとしている。
もしくはそもそもこの実験では129/SvのCAGホモは使われていなくて実際には129/Sv carrying Rosa26-gfp だったのに若山さんが後から嘘にしているのかもしれない。この場合は理研も嘘に加担していることになる。というのも笹井さんや丹羽さんは129/Sv carrying Rosa26-gfp の記述に何の疑念も感じていません。理研にはそういうマウスはいろいろとあるし、丹羽さんも実験で使っている。ところが桂報告書は、「129/Sv carrying Rosa26-gfp マウスは理研 CDB に導入された記録や飼育記録はないことから、 これは誤記と考えられ、若山氏の説明によればここで言及された STAP 幹細胞は AC129 であった可能性が高い。」と書いている。
これは入手経路にマウス会社との裏ルートがある場合は本当のことは書けません。裏ルートというのは独法にありがちな余った予算を関連会社に商品を買ったことにして預ける。民間でもよく行われて国税に摘発される架空取引です。役所には税務署は入らない。これで予算を余らせないで済む。まあ、そういうところは蛇の道は蛇で、誰でも知ってるが、そんな話はここでの目的ではありません。
③④に関してはそもそもどんなマウスを小保方さんに渡したかはどこにも書かれていない。ただ、FLS-Tと書かれているから、FLSは「僕のマウス」を渡したと言ってるんだから、この場合もそうだろうと推測されているだけです。TはTeruで、若山さんの名前の照彦の略です。若山さんは一人で幹細胞まで樹立してそれを山梨大に持ち去ったわけですが、無論これは一人でできるかと試したものなので小保方さんには残していない。そして桂調査とBCA調査で129B6F1 ES1で捏造されていると証明されて、小保方さんが「僕のマウス」ESをインキュベーターの中でそっとコンタミしたのだという結論にさせようとしたのに、どれがどれと分からないと思っていたからでしょうが、ドジなことに若山さんは大本の129B6F1 ES1を山梨大に持ち去っていた。そして小保方さんのフリーザーボックスに129B6F1 ES6が残されていて、小保方さんは桂報告の公表前にこれに関して由来不明と答えているんです。警察だったらまず最初に疑われるのは若山さんです。
AC129の実験を行うとき若山さんはAC129のCAGホモを渡したと言ってるわけです。桂報告書には何らその証拠は提示されていません。FLSのときに「僕のマウス」を渡したという話も何の証拠も提示されていません。実験ノート記載くらいは見たんでしょうか。実験ノートにも書かれていなかったようですが。ただ若山さんがそういってるということです。呆れます。ノートに書かれているのなら実物を提示してくれないと困りますが、Ooboeさんのパートナー氏やDORAさんが開示請求しても応じません。以下はDORAさん資料です。

小保方さんが米国側の要請で開示できないということはさんざん非難したくせに。そして小保方さんが入院中に手元に置いていた実験ノートは強引に持ち帰り、全部コピーを取って、しかもNHKにその全コピーを違法に流出させた。何をやってるのか。何しろ今までごまかしばかりやってるから、彼らのやることは何事も鵜呑みにはできないと世間は見てます。
ただ、129/Svは白毛ですから小保方さんは白いのを受け取ったとは確認してます。若山さんはこの時キメラは作らなかったと証言してますが、小保方さんは作られたと論文に書いている。よくもまあ、こんなに食い違うものだね。若山さんは論文読んでないのでしょうか。
c.AC129実験の目的問題はAC129の実験とは何なのかということです。再掲しましょう。
>>
(d) STAP 幹細胞 AC129 は、129B6F1 マウスから作製された受精卵 ES 細胞に由来する
(調査結果) STAP 幹細胞株樹立に遺伝的背景が及ぼす影響を調べる実験が、若山氏により 2012 年 夏から秋にかけて行われ、2012 年 9 月 4 日に STAP 幹細胞 AC129-1 および AC129-2 が樹 立された。若山氏が交配した 129/Sv-CAG-GFP マウス(CAG-GFP 遺伝子が第 18 染色体に挿 入されたホモ接合体)由来の脾臓 CD45 陽性細胞を材料とし、小保方氏が STAP 細胞を作 製し、それを用いて若山氏が STAP 幹細胞として AC129 を樹立した。この細胞ストック はCDB若山研が山梨大へ移転する際に山梨大へ運ばれ、また一部は小保方研へ分与され、 それぞれフリーザーに保管されていた。このうち、小保方研フリーザーに保管されてい た STAP 幹細胞 AC129-1 について、SNPs マーカーの TaqMan PCR 法による解析を行い、さ らに NGS により全ゲノム DNA 配列を解析した。同じく STAP 幹細胞 FLS の対照として CAG-GFP マウスから作製された受精卵 ES 細胞(129B6 F1ES)の解析も同時に行った。得 られたデータを他の細胞等の解析結果や公開データと照合した結果、以下のことが判明 した。 「同じく STAP 幹細胞 FLS の対照として CAG-GFP マウスから作製された受精卵 ES 細胞(129B6 F1ES)の解析も同時に行った。」という書き方がレトリカルであるということがわかります。AC129-1が全解析されて、続けて「同じく」と書くとそちらも全解析されたかのように誤読されやすいのを分かってこう書いているんです。実際にはどういう解析なのか示されていないが、先に考察した通りです。示されているのは129B6 F1ES6の全解析結果だけなのに、それを含めて「(129B6 F1ES)の解析も同時に行った。」とラインをわざと書かずに紛らわしくさせているのです。報告書のスライド版も同じです。全解析したもののみのリストであるはずなのに129B6F1とだけ書かれていて、6つある中のES-6であってES-1ではないということが示されていない。意図的な隠蔽です。
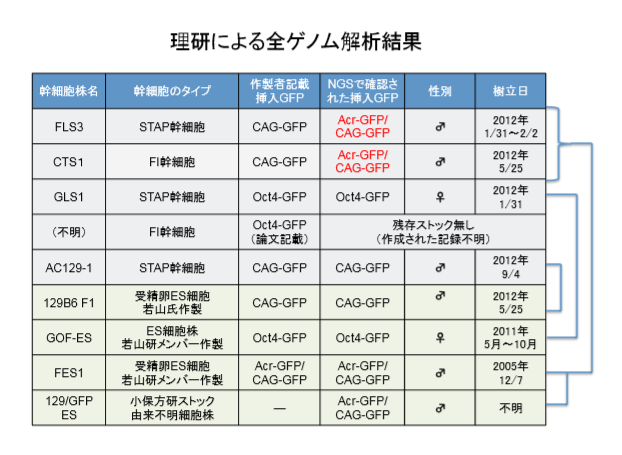
ともあれAC129の実験の目的は「STAP 幹細胞株樹立に遺伝的背景が及ぼす影響」だということです。
2012年の8月にもなってまだ「STAP 幹細胞株樹立に遺伝的背景が及ぼす影響を調べる実験」をしていたというんですが、2011年に既にF1とGOFで幹細胞化させています。最初のF1は「僕のマウス」ではないことは桂報告書が明らかにしている。翌年の初頭に又「僕のマウス」とGOFでやってる。B6GFP x DBA/2もやってます。その後はTCRの実験で又STAP細胞を作ってます。この時にキメラや幹細胞は以前のを使って、新たには作らなかったのでしょうか。更に胎盤が光ったという実験でSTAP細胞を作らせてキメラを作ってます。そのマウスの背景は何なんでしょうか。胎盤が光ってたんで最低でもCAGです。この時もSTAP細胞はリンパ球を使ってるはずです。コントロールの受精卵ESでもキメラを作って胎盤を小保方さんに渡しました。念のためにもう一回は再掲して置きましょう。
若山さんはこの胎盤実験に関して左右の実験を一人で同時期に行ってることになるんです。これをやって酸浴リンパ球のナイフカットによるキメラ胚移植と、コントロールES細胞のキメラ胚移植で、細胞の大きさと、自分で引き延ばすパイプの太さが違うということに気づかない人はいません。>>
(全録)STAP細胞論文の共著者・若山照彦教授会見 質疑応答(2/4)
8:30/42:12~
(朝日新聞岡崎女性記者)
で、先生は実際に、その、STAP細胞からキメラマウスを造ったり、その、幹細胞を作成されたりして、実際に、その、細胞を見てるわけですけど、見た目、外観からすると、全く今までと違った細胞で、あの、他の類似した細胞というのは見たことなかったのでしょうか。
(若山さん)
マイクロマニピュレーターの上に細胞を乗せてしまって、そこからキメラとかを作るわけですが、その状態になってしまうと、あと、あの、今回に関しては、それまでずっと失敗続きだったということもあって、ええ、普通、キメラマウスを作るときは、細胞をバラバラにして、キメラを作るんですが、塊のまま入れてみようという、そういうアイデアで実験をしたということもあり、いつもと違う手法を取り入れた、その時に成功したんです。そのために、その時の細胞が以前と違っていたかどうかというのが分からないままなんです。
(朝日新聞岡崎女性記者)
では、その、もしかしたら、その、万が一ES細胞だったとした場合、それを、これが新しい細胞ですと言われて、ほんとに、見分けがつかなかったんですかね。
(若山さん)
そうですね。その時点では、あの、新しい手法でやってしまったので、見慣れた外観は全くないので、どの細胞だったかということは区別できなかったと思います。嘘をついています。分からないことはありません。笹井さんも分かるはずだと言いました。若山さんはナイフカットは最初に一回やって写真撮っただけなんです。嘘ついてるんです。まあ、もはや我々はそれは前提でAC129の問題も考えています。
- 2019/08/27(火) 10:12:38|
- AC129
-
-
| コメント:0
『AC129を巡る問題』(1) AC129とは何であったのかA.経緯AC129-1,2とFLS-1,2は129B6F1ES-1によって捏造されたと桂報告及びBCA報告は主張している。
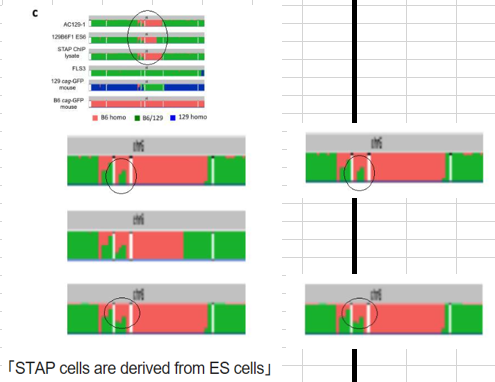
BCA報告のその部分は以下です。(参考 *ttps://www.researchgate.net/publication/283439572_STAP_cells_are_derived_from_ES_cells)
>>
Control genomic DNA sequences for STAP cell chromatin immunoprecipitation sequencing (ChIP-seq) experiments (Fig. 4 in ref. 2) had been deposited in the NCBI database(2). To gain sufficient sequencing coverage, we re-sequenced the genomic DNA prepared from the STAP cell lysate used for ChIP-seq (Extended Data Fig. 1a). We confirmed that this STAP cell sample shared all the genomic characteristics described above for 129B6F1 ES1 (Extended Data Fig. 2c),indicating that the STAP cell sample used for ChIP-seq was derived from 129B6F1 ES1 cells.ここで(Fig. 4 in ref. 2)と指定されているのは、Letter Figure 4-bのことです。
その図のSTAPとESのデータが入れ替わっているというのが若山さんによるリトラクション理由書にあるのはよく知られている。序ですから引用しておきましょう。
>>
(4) In Fig. 4b of the Letter, STAP cell and ES cell are wrongly labelled in a reverse manner.Letter Figure 4-bは以下です。よくこの程度の粗雑な簡略図でSTAPとESの取り違えに気づけたものです。誰かラボ内に手伝った人でもいて、元データを持っていた人がいたことが推察されます。
BCA報告の(Extended Data Fig. 2c)の図は以下です。
上表のSTAP ChIP lysateに関してBCA報告本文に説明がありません。書かれているのはNCBIのデータベースと、自分たちの分析図です。
>>
Control genomic DNA sequences for STAP cell chromatin immunoprecipitation sequencing (ChIP-seq) experiments (Fig. 4 in ref. 2) had been deposited in the NCBI database(2). To gain sufficient sequencing coverage, we re-sequenced the genomic DNA prepared from the STAP cell lysate used for ChIP-seq (Extended Data Fig. 1a). NCBIのデータベースに関しては、すでに我々が以前分類しているもので以下の分です。
>>
ChIP-Seq
⑩SRR1171582 小保方 Low pH treated CD45 positive Cells:ChIPSeq.H3K27me3 Oct3/4 expressing cells C57BL/6x129/Sv 129xB6-CAGヘテロ
⑪SRR1171583 小保方 Low pH treated CD45 positive Cells:ChIPSeq.H3K4me3 Oct3/4 expressing cells C57BL/6x129/Sv 129xB6-CAGヘテロ
⑫SRR1171584 小保方 Low pH treated CD45 positive Cells:ChIPSeq.input Oct3/4 expressing cells C57BL/6x129/Sv 129xB6-CAGヘテロこれがLetter Figure 4-bのSTAPの欄で上の緑がH3K4me3で下の赤がH3K27me3になっています。若山さんの取り消し理由書に書かれているのはESの分とSTAPの分が逆に提示されているという主張です。このデータはTs.Marker氏が再分析していますが、素人では見方が分からなくて入れ替わっているのかどうか確認できません。そのことに関する彼の見解もありません。
他方、⑫に関してデータだけでなく、混合液であるSTAP ChIP lysateが残っていたのは別の話ですから由来を説明しないと分かりませんが、BCA報告の(Extended Data Fig. 2c)の図は全く同じものが桂報告書のスライド版にも使われていて、本文16Pに以下のようにあるが、どういう形で保管されていたのかは説明されていない。
>>
3)STAP 細胞由来 ChIP-seq (input)サンプルは 129B6 F1ES1 から取得された
(調査結果) 小保方氏が CDB ゲノム資源解析ユニット(以下「GRAS」という)に持参し残されてい た STAP 細胞由来 ChIP-seq (input)サンプルを再度 NGS 解析した結果、STAP 細胞由来と される ChIP-seq input データは CAG-GFP の挿入を持つ 129xB6 へテロ系統由来の細胞か ら取得されたものと判明した。さらに SNPs の解析、特異的な欠失変異の解析により(2 -3-1-1(d)参照のこと)CAG-GFP が挿入された 129B6 F1ES1 とほぼ同一細胞由来の データであることが明らかとなった。2年前の生データがSTAP細胞だけ冷凍されて残っているというのも変で、残っているならこの時の試料の全てのはずですが、記者会見では伊藤氏が口頭で記者の質問に答えて、この時の分析の試料がGRASにたまたま残っていたのだと証言しただけで、他の試料の有無に関しての更なる追及も無かった。又その保管に関して文書で追加説明があったということもない。伊藤さんはBCA報告の著者の一人です。
>>
Daijiro Konno, Takeya Kasukawa, Kosuke Hashimoto, Takehiko Itoh, Taeko Suetsugu, Ikuo Miura, Shigeharu Wakana, Piero Carninci & Fumio Matsuzaki桂会見での古田女史の質問に対する答えは以下でした。言い終わった後の彼のしぐさと表情は精神分析学の研究対象に好例かもしれない。(*ttps://www.youtube.com/watch?v=M9oJGioHvIQ 1:15:00から)
>>
ただ、唯一やったのが、あ、GRASの方に残っていたChIP seq のinput 、しかもSTAP細胞のインプットはDNAとして残っていたので、それが10ページバワーポイントの、えーと、ところに書いてある、3番のアイテマイズして書いてあるところですけれども、これに関しては、えーと、30倍になるだけ、他のゲノムと同量となるまで読んでいます。そして、読んだ結果、この、同一であるということを、ま、認定したということで、それ以外に、それ以外に関しては、ま、残っていませんのでやってない、やってません。なぜ、たくさんある中の、唯一、STAP ChIP lysateだけが残されていたのか、どういう形で唯一残されていたのか、誰かの恣意で残されていたのか、それともGRASには残し方に関する規則は無いのか。裁判だったらまず最初に検討されるもので、ここにちゃんとした答えが無いと証拠採用はされない。
B.報告書の非論理と重大な疑念a.非論理まず、先にBCA報告のこの件に関するあからさまな嘘を指摘しておきましょう。
>>
We confirmed that this STAP cell sample shared all the genomic characteristics described above for 129B6F1 ES1 (Extended Data Fig. 2c),indicating that the STAP cell sample used for ChIP-seq was derived from 129B6F1 ES1 cells.上に貼り付けた(Extended Data Fig. 2c)には
129B6F1 ES1は存在していません。あるのはES6です。ES6を図示していながら、これもどこから持ってきたかも正式に記述しないSTAP ChIP lysateが、図に無いES1と同じSNPsパターンであることを確認したと書いているわけです。
東の空を指さしながら美しい夕焼けですねと言ってるような論理です。言ってる事実が少なくとも指さしている方向には無いので、言ってる事自体も本当か嘘か分からない。科学者が書いたとはとても思えない
非論理な文章です。
b.最大の疑念以下に示すのは木星リストですが、まず129B6F1 ES1はそもそも小保方フリーザーには存在していないものです。右端の書き込みは調査のために管理部署から持ち出された記録です。129 B6 ES-4と5には持ち出された記録がないということにも注意が必要でしょうね。PCRに掛けられたデータがありますから持ち出したことは確かのはずですが、記録をつけないまま杜撰に持ち出して調べている実態がうかがわれる


では129B6F1 ES1はどこにあったのか。以下の若山さんの事後MTA契約書添付の持ち出しリストの中にある。下から二段目です。

よりによって若山さんは129B6F1GFP-1を持ち出していました。AC129-1,2もFLS-T 1,2も若山さんが持ち出していたControl ES cell の 129B6F1GFP-1 で捏造されているとBCA報告は証明しているわけです。ところが桂報告書もBCA報告書もAC129とFLS-Tは129B6F1ES1によって捏造されたと声高に主張しながら、その試料がどこにあったかを一切書いてない。小保方さんのフリーザーにあったものではないどころか
若山さんが山梨大に持ち出していたのです。桂報告書もBCA報告も、AC129の実験は小保方さんが若山さんに129B6F1GFP-1を渡して捏造させたものと示唆しているのですが、使われた細胞は理研には無くて、山梨大にあったのです。そして、報告書はこんな重要な事実を報告していないのです。これが分かったのは木星さんを中心にした大勢の人々の公開資料の取り寄せ努力によるもので、理研自らが発表したことではありません。むしろ
彼らはそれを隠していたことになるのです。これが疑念の最初で最大のものです。
c.その他の疑念次に、桂報告とBCA報告の遺伝子解析によるAC129と129B6F1ES-1との一致証明に関する疑義です。これは以下の表で、桂報告書とBCA報告のどちらにも掲載されています。BCAのは遺伝子異常のPCR結果もついていますからそれを貼り付けます。
この表にある129B6F1 ES4と5には持ち出された記録がないということと、ES1が山梨の若山さんのフリーザーにあったということが書かれていないということは既に指摘した通りです。通常の事件ですと、こういう情報が揃うとまず最初に疑われなければならないのは小保方さんではなくて若山さんの方なんですけどね。先にこの所謂「僕のマウス」ESのサンプル保管の状況から検討しましょう。
(サンプルの状況)これに関しては既に見てきた通り二通りのラベル表記がある。若山さんの持ち出しリストに嘘か誠か6ライン樹立したと書かれています。嘘か誠かというのは、TSに関しては2ラインになってるが実際には129 B6 TS-3,4,5,6が理研側に残されていますからこれも6ラインのようです。嘘か勘違いかは問わず、今はESに関してのみ考えましょう。想定されるもともとあったラインは以下です。
①129 B6 ES-1,2,3,4,5,6
②129B6F1 GFP ES-1,2,3,4,5,6
ここで重要なのは今桂報告として知られている調査結果が世間に知られる以前の聞き取り調査段階で、小保方さんが①は若山さんが作ったと証言しているのに、離れた場所に入っていた②の6に関して「由来不明」と証言していることです。
つまり、小保方さんが知っているラベルの書き方は①であったということです。そして、②の1は若山さんが持っていて、②の6が誰かの置忘れか、意図的にそこに置かれたものだということになる。現状は以下です。
>>
ES細胞 129B6F1 GFP-1(若山持ち出しリスト)
129 B6 ES-2(小保方さん「若山TL樹立」証言)
129 B6 ES-3(小保方さん「若山TL樹立」証言)
129 B6 ES-4(小保方さん「若山TL樹立」証言)
129 B6 ES-5(小保方さん「若山TL樹立」証言)
129B6F1 GFP ES-6(小保方さん「由来不明」証言)
2013年の3月の引っ越し時に若山さんによるサンプルの整理が行われている。そして山梨大から放医研にサンプル調査に出したのは1年後の2014年の3月です。そして4月になってMTAの持ち出しリストに自分は現在1しか持ってないように書いた。
彼は記者会見で放医研に出した129B6F1 ESを5株と言っています。なぜ6株では無いのか。
そして、この5株は放医研側のPCR結果のデータには1,2,3,4,6と書かれている。5がなぜ提出されなかったのかの理由説明もない。
更にMTAの持ち出しリストには6株樹立されて、今自分が持っているのは1だけのように記載されている。放医研には株分けして出したのであれば少なくとも1,2,3,4,6が持ち出しリスト上に無ければならない。仮に株分けせず手持ちのものをそのまま出したのなら、放医研に出したものは記載しないのなら何もないことになる。1だけがあるという理由は1だけは株分け分があったということになるしかない。
ところがこのESは理研側でも調査されていているからにはそのときには全ラインあったということになる。理研側の調査したサンプルの由来はどうなっているのか。
小保方さんのフリーザーに残されていたものとその松崎氏の持ち出し状況は以下です。
>>
129 B6 ES-2 (2014/6/20 松崎氏持ち出し記録あり)
129 B6 ES-3 (2014/6/20 松崎氏持ち出し記録あり)
129 B6 ES-4
129 B6 ES-5
129B6F1GFP ES-6(2014/6/9 松崎氏持ち出し記録あり)
若山さんの記者会見スライドにあった放医研の記載の仕方は以下でした。ラベルは全部新たに書き換えられて提出されていることが分かります。
>>
129B6F1-1 ES
129B6F1-2 ES
129B6F1-3 ES
129B6F1-4 ES
129B6F1-6 ES
放医研のサンプルを回して貰ったのなら5がありませんが、どこから持ってきたのでしょうかね。129 B6 ES-5 は少なくとも持ち出し管理記録上は持ち出されていないことになっている。調査が如何に杜撰で恣意的かということです。
ただし、1~6は欠失重複等が全部違っていますから理研側が6種類揃えたことは間違いありません。何かの中身が重複しているということはありません。調査細胞の入手経緯が明らかになっていないだけです。
我々の推測では[129B6F1 GFP ES-6(小保方さん「由来不明」証言)]を小保方さんのボックスに置いたのは若山さんだと推測しています。若山さんは1を置いておいても6を置いておいても大して違わないと思っていた。というのも遺伝子異常があって全ライン識別されるとは思ってもいなかったでしょうし、雌雄の違いも考えている暇がなかったんでしょうね。1も6もたまたま雄でしたが、違ってたら言い訳のつかないことになってたでしょう。核型解析に出せばすぐ分かりますから事前に雌雄確認をしていたかもしれません。ただ、遺伝子異常まではさしもの若山さんも気づかなかったでしょう。ES-1は無論、直前にFLS-T1,T2を作っていた時に使った株分け分が手元に残ったのではないか。これが致命傷になった。これ見よがしに置かれていたES-6は使われていなくて、若山さんが山梨に持ち出していたES-1が使われていたという藪蛇になってしまった。
問題は理研側に残されていた分で129 B6 ES-1と129 B6 ES-6がなぜ無いのかということです。このコントロールESは小保方さんは論文に使っていますから、実際に使用している。別名でどこかにあるかもしれません。後に詳しく調べてみましょう。
(SNPs解析)SNPsでAC129タイプと書かれているのはES1タイプということです。ES1タイプのマウスとES6タイプの2タイプの卵が使われているということです。なぜこういうことになるのかを考えてみる。
SNPは一塩基性多型と呼ばれている突然変異で人間でもそれぞれの人の顔が違うというような表現型の多様性等を作っているものですが、無論人間であることは共通ですから大半の遺伝子は共通です。
一方近交系マウスは20世代以上の兄妹交配を重ねて相同染色体の同一遺伝子座の遺伝子をホモにそろえてありますからSNPsもホモです。近交系マウスはマウス業者(ラボ)が作って系統立てて管理しているわけですが、例えば129/SvJ独特のSNPs分布というのは最初にジャクソン研究所で作ったマウスのSNPsを引き継いでいるものです。B6のラインを作るときは又別のマウスを使いますからこれは亜種が違うので当然全然違う分布になる。しかし、129でもB6でも最初に使われたマウスの個性が引き継がれるので仮に別の人が一から作ると別のSNPs分布になる。それでは研究目的に使えないので、同じ名前にはなりません。ですから例えば129/SvJと言ったら最初にジャクソン研究所の作ったマウスと決まっていてそれが国際的にも引き継がれていて、SNPs分布も決まっている。
継代繁殖で新たにできるSNPsというのはわずかなので、長く蓄積すると更に別の表記を加えますが、直接的には関係ないんです。
で、SNPsパターンでES1タイプとES6タイプの二つあるというのは、雑種交配させたんですから、129とB6の混ざり方の違う2個の受精卵が使われているということになる。完全な近交系マウスでは減数分裂時にできてくる卵も精子もそれぞれに遺伝子配列は全く同じですからこんなことになることはない。従って一方が完全な近交系マウスになってないということです。その原因は所謂「僕のマウス」作成の経緯にある。
若山さんはロックフェラー大学にいるときにB6のCAGホモマウスを作った。日本に戻ってからそれを129と掛け合わせてその後戻し交配を続けてGFPだけを129に移し替えた。129もB6も無論近交系で、マウス生産業者が20世代以上の兄妹交配を続けて対立遺伝子をホモにそろえて販売しているものです。契約があって原則自家繁殖させてはいけない。特に人にあげてはいけないです。マウス業者がその純度を保証できないから予期せぬところでの責任を負いきれないというのが理由です。
最初にB6の卵にウイルスベクターを使ってCAG-GFPを挿入する。どこに入ったかは分からないが、入ったら生まれたマウスは蛍光する。この蛍光している一匹のマウスに又B6を掛け合わせてたくさん子供を作る。この時の子マウスのCAG-GFPが光っている雄雌だけをメイティングする。GFPヘテロ同士の掛け合わせになります。ヘテロ同士を掛け合わせると第3世代で1/4の確率でホモができる。ランダムに交配させて十分な子マウスを作って一匹もGFP無しの子マウスが生まれてこなかったペアはホモ同士という確認で継代を重ねることを省略できる。もともとが近交系マウスですからGFP以外は全部ホモです。そういうことをしないで念を入れて最初から20世代継代してもホモになりますね。(放医研の検査で18番にホモに入っていました。)
それに対して129に移し替えるというのはよりテクニカルです。ただ単に129のCAGマウスを作るだけならB6と同じやり方でできる。でもこれだと又GFPがどこに入るか分からない。GFPが染色体の同じ場所にホモで入っているB6と129が作りたいということです。
それでまず一匹のB6-CAGホモマウスと一匹の普通の129とを交配させた。これは最初129B6F1です。B6がGFPホモですから子供には全部GFPがヘテロに来る。ただし、雑種にしましたから他の遺伝子もヘテロに変わってしまっています。
そこで129への戻し交配を行って、GFP遺伝子以外を親の129の近交系に戻す。そのやり方はGFP蛍光のある子供に対してのみ戻し交配を行うんです。これを繰り返すとGFPのみヘテロの129近交系マウスに戻ります。このヘテロ同士を掛け合わせて129GFPホモを作るんですが、この戻し交配が完全に行われていないとGFP以外の他の遺伝子にもB6の遺伝子が残ってしまう。それが上に貼り付けているBCA報告のExtended Data Figure 2-cのSNPs分析です。再掲しておきましょう。
上図のリジェンドです。
>>
c, Distribution of B6-type and 129-type SNPs along chromosome 6. The B6-homozygous SNP cluster(magenta) in the middle, which probably arose from the inheritance of the parental 129, is heterogeneous in length among six control ES cell lines. The four cag-GFP STAP stem-cell lines share the same length of the B6 SNP cluster with control ES 129B6F1 ES1. Note that the 129/B6-heterozygous SNP region in the 129 cag-GFP mouse is longer than that of AC129-1.下二つが今説明した親マウスの6番染色体のSNPs分布図です。B6は全部ピンク(magenta) です。これはロックフェラー大学で作ったGFPマウスです。GFPを挿入しただけですから対立遺伝子は全く同じで全部ホモにそろっていますから問題ない。対して129の方は全部ブルーになってないといけませんが、ピンクの部分とグリーンの部分が残ってます。ピンクの部分は相同染色体の両方がB6で、グリーンは一方がB6で他方が129ということです。戻し交配の回数が不足している状態でGFPヘテロ同士を掛け合わせてホモにしてしまった結果かもしれません。因みにGFPホモであるかどうかは更にもう一代子供を作って、全部蛍光しているかどうかで確認できます。
問題は129です。近交系マウスに戻っていない。厳密にいうとGFPだけでなく、129/Svだということすらできない状態です。もう少し長くバッククロスを続けてから兄妹交配させないといけなかったのかもしれない。でも、調べない限り本人は気づいていませんからやり直すことはできない。できたものの兄妹交配を続けるだけです。しかし、これはこれなりに20世代交配させていたら近交系マウスになります。
マウスは20日で生まれて更に妊娠可能になるまで50日かかります。70日の20継代は1400日です。3.8年です。若山さんは日本に帰ってきてから129へのGFP移し替えを行いましたから、戻し交配でも3.8年かかっている。しかし、その後の20継代維持を入れると計7.6年ですが、理研の10年目だったわけですから、ちゃんと継代できていたら、全部ホモにそろっていたはずです。にもかかわらずヘテロ領域がこんなにあるというのはもっと近い時期にB6とのコンタミがあったということになるのではないでしょうか。又そもそもの129には歴史的にジャクソン研究所で一度B6が混入したという事件がありました。従ってピンク部分はもともと存在していたものかもしれません。
そもそも論としてこういう図形がどうしてできるかということですが、ひとつづつの対立遺伝子のあり方はホモかヘテロしかありません。大多数のDNAはマウスである限り共通ですから、129かB6のどちらということはできません。言えるのはSNPのある個所だけです。
このケースの場合で、6番染色体のDNAの並びをある一定区間で区切って1単位とします。この長さは適当に人が決める。この区間にSNPsが何もなければ白抜きで表示する。遺伝子のあるところは大事なところなんでSNPsは少ない。そういう場所は白抜きになる。存在しているSNPsがB6特異的な変異だったらピンクで表示する。129特異的な変異だったらブルーで表示する。半々だったらグリーンです。F1ですから父と母とから来た両方があるのが普通です。だから大半グリーンになる。
では混じっているが半々でないときはどうするか。半々の部分とB6が勝ってる部分、129が勝っている部分と1単位の中で棒グラフとして表示している。
1単位の区間を細かくすると例えば以下のBCA報告のExtended Data Figure 1-c図のようになるし、区間を広くとると上のExtended Data Fig. 2cのようなものになるわけです。
ともあれ、母親の129の染色体の一部にヘテロ部分があったわけですから卵原細胞が減数分裂するとき別々のSNPs分布を持つ二種類の卵ができる。マウスは多産ですのでたくさんの受精卵ができて、胚盤胞にまで成長させてから中のインナーセルマスを取り出してES細胞を培養する。8株できたわけです。それがES1型とES6型に分かれたのはそういう理由です。雌雄は以下です。
>>
ES1タイプ→ES1(オス)、ES2(メス)、ES5(オス)
ES6タイプ→ES3(オス)、ES4(メス)、ES6(オス)
この解析結果の不可思議はES6以外のSNPs解析データが図示されていないことです。このことは後に述べます。
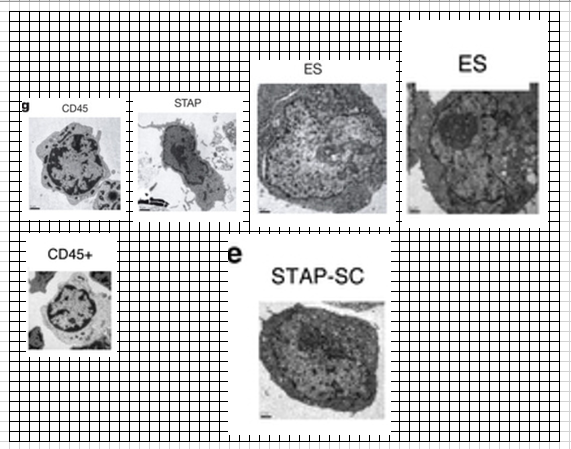
(関連細胞)Letter Figure 4-bで使われている各種細胞は以下です。
>>
①CD45
②STAP
③ES
④STAP-SC
⑤FI-SC
⑥TS
Letter Figure 4-bそのものはこれです。再掲します。
この図に対応しているのが公共データベースに登録されている以下のデータです。
>>
ChIP-Seq
①SRR1171553 小保方 CD45 positive Cells:ChIPSeq.H3K27me3 derived from spleen Oct3/4::gfp C57BL/6 GOF
②SRR1171554 小保方 CD45 positive Cells:ChIPSeq.H3K4me3 derived from spleen Oct3/4::gfp C57BL/6 GOF
③SRR1171555 小保方 CD45 positive Cells:ChIPSeq.input derived from spleen Oct3/4::gfp C57BL/6 GOF
④SRR1171562 若山 Embryonic Stem Cells:ChIPSeq.H3K27me3 C57BL/6x129/Sv
⑤SRR1171563 若山 Embryonic Stem Cells:ChIPSeq.H3K4me3 C57BL/6x129/Sv
⑥SRR1171564 若山 Embryonic Stem Cells:ChIPSeq.input C57BL/6x129/Sv
⑦SRR1171567 若山 FI-SC(Fgf Induced Stem Cells):ChIPSeq.H3K27me3 Oct3/4 expressing cells C57BL/6x129/Sv 129xB6-Acr-CAG
⑧SRR1171568 若山 FI-SC(Fgf Induced Stem Cells):ChIPSeq.H3K4me3 Oct3/4 expressing cells C57BL/6x129/Sv 129xB6-Acr-CAG
⑨SRR1171569 若山 FI-SC(Fgf Induced Stem Cells):ChIPSeq.input Oct3/4 expressing cells C57BL/6x129/Sv 129xB6-Acr-CAG
⑩SRR1171582 小保方 Low pH treated CD45 positive Cells:ChIPSeq.H3K27me3 Oct3/4 expressing cells C57BL/6x129/Sv 129xB6-CAG
⑪SRR1171583 小保方 Low pH treated CD45 positive Cells:ChIPSeq.H3K4me3 Oct3/4 expressing cells C57BL/6x129/Sv 129xB6-CAG
⑫SRR1171584 小保方 Low pH treated CD45 positive Cells:ChIPSeq.input Oct3/4 expressing cells C57BL/6x129/Sv 129xB6-CAG
⑬SRR1171587 若山 STAP-SC(STAP derived Stem Cells):ChIPSeq.H3K27me3 Oct3/4 expressing cells C57BL/6x129/Sv 129xB6-CAG
⑭SRR1171588 若山 STAP-SC(STAP derived Stem Cells):ChIPSeq.H3K4me3 Oct3/4 expressing cells C57BL/6x129/Sv 129xB6-CAG
⑮SRR1171589 若山 STAP-SC(STAP derived Stem Cells):ChIPSeq.input Oct3/4 expressing cells C57BL/6x129/Sv 129xB6-CAG
⑯SRR1171592 丹羽研 Trophoblast Stem Cells:ChIPSeq.H3K27me3 CD1
⑰SRR1171593 丹羽研 Trophoblast Stem Cells:ChIPSeq.H3K4me3 CD1
⑱SRR1171594 丹羽研 Trophoblast Stem Cells:ChIPSeq.input CD1最初に注意しておかなければなりませんが、これらの試料がGRASに提出されたのは2012年の8月です。従って細胞の製作者は基本小保方さんと若山さんです。8月の解析は笹井さん参加前の実験結果で、しかも幹細胞化実験ですから、3誌論文の中には取り入れられてないものです。手記によると8月になってから若山さんが幹細胞化実験の成果を自分に51%割合で特許申請手続きを開始したとされている。そこから米国側と揉めて11月には小保方さんが米国に引き上げようとした経緯につながっている。
笹井さんが引き継いでからNGS解析をつけようということになってこの8月のデータが検討された。ChIP seqのデータもこのときにLetter Figure 4-bに纏められた。
若山さんは多忙で手伝えないということですから、新たに幹細胞やキメラを作ることはできない。公開データは笹井さん参加後にレター論文に使われたものです。若山さんは手伝っていませんから、若山さんが作ったものは2012年8月以前のものです。ES細胞とFI幹細胞は若山さんしか作れない。STAP幹細胞は小保方さんがSTAP細胞を作って若山さんがそれを使って作成した。CD45細胞とSTAP細胞は小保方さんはマウスさえあれば何時でも作れる。TS細胞は若山さんしか作れませんが、若山さんはF1で作ったと言ってますから、CD1マウス背景のTSは丹羽研のものと分かる。このことは若山さんの作ったTSが分化しているようだったので丹羽さんが提供したと桂報告書にも書かれています。桂報告書26P。
>>
TSについては細胞培養時に分化し たことが考えられたことから、丹羽研のメンバーが培養・サンプル調製を行ったものを 追加して作図に用いた。⑯⑰⑱は丹羽研のものと分かっているわけです。論文の論旨としては比較のためのコントロールですから誰のTSでもいい。2012年8月に提出されていたもともとの細胞はF1背景のTS細胞だった。
①②③は白血球ですが、GOFだと書かれている。これも論文ではGOFとF1が使われているわけですが、どちらでもいい。ただし、2012年8月のGRAS提出目的はレター論文用ではありません。笹井さんはまだ参加してない時期です。特許申請手続きを始めていますから、若山さん主導の幹細胞論文や特許申請書に添付するデータとして使う目的です。この時にGOFであったかどうかは分かりません。もしかしたら他と同じF1の分であったかもしれない。
その他は2012年8月のときのままです。酸浴細胞は小保方さんは何時でも作れますが、若山研のF1マウスは笹井研時代には使えませんから当時のままだと分かる。そしてすべての試料はF1背景です。ただし、理研が2012/2/13にNCBIに提出したときには今見られるとおりのC57BL/6x129/Svになっていた。しかし、小保方さんが理研に提出した2012/11/3の時点でどう書かれていたかは分かっていません。ここには若山さんの作ったES細胞があってそのマウス背景は129/SvxC57BL/6だということは分かっていますから、この登録の少なくともこの分は間違っている。ただ、それを小保方さんが間違えたのかは分かりません。
小保方さんはNCBIに直接登録はしていない。行ったのは理研の別人です。ここではF1であるという事にとどめておきます。
④⑤⑥がそのES細胞です。
⑦⑧⑨はFI幹細胞ですが、後の調査でアクロシンが出た。桂報告では太田ESだということになっていますが、若山さんは細胞の大きさで気づかないということはあり得ないと証明済ですので、これは岡部マウスとのF1由来の小保方細胞核使用ntESをTS培地誘導したものと分かる。
⑩⑪⑫がSTAP細胞ですが、これはアクロシンが出ていない。しかもCAG-GFPです。129でもB6でもCAG-GFPの入っているマウスは「僕のマウス」の親しか知られていない。小保方さんは見かけでは親の雌雄とGFPがホモかヘテロかはわかりません。
⑬⑭⑮が上の⑩⑪⑫をSTAP幹細胞誘導したものということになるが、これも後の調査でCAG-GFPと判明している。我々の判断ではこれもntES化しているということになるが、胎盤が光ると言われたキメラは小保方さんはB6/129としていますから、その時にSTAP幹細胞が作られていると知らされているのかもしれない。いずれにせよ。FLSは「僕のマウス」を渡したが中身は太田ESだったとされているが、
ここに公共データ登録では背景がB6x129と記載されている細胞で、事実中身が「僕のマウス」から作られているSTAP幹細胞と称する細胞があるということは驚きで、このことに誰も気づいていません。もしこれが「僕のマウス」ESの1~6のどれかなのなら、今でもデータはあるのですから調べたらわかります。しかし、SNPs解析も遺伝子異常解析もされていません。ただ、マウス背景が129B6F1のCAGホモだと桂報告が言ってるだけです。
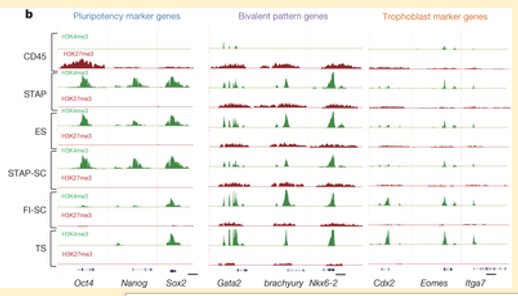
Letter Figure 4-bは、ES細胞に代表される多能性遺伝子(Pluripotency marker genes)であるOct4,Nanog,Sox2、二価クロマチン(Bivatent pattern genes)であるGata2,brachyury,Nkx6-2、そしてTS細胞特異的遺伝子(Trophoblast maker genes)であるCdx2,Enomes Itgα7に関して、それぞれの、その巻き取られているヒストン領域がH3K4me3(H3ヒストンのK4サイトのリジンの3つあるアミノ基が全部メチル基に置き換わっているトリメチル化と呼ばれる現象を起こしている)とH3K27me3の状態にあるDNAをクロマチン免疫沈降(Chip=chromatin immunoprecipitation)で抽出した後に定量PCRに掛けた結果である。
(H3K27me3)は常時転写抑制として働き、(H3K4me3)は活性化を許可する働きを持つ。二価クロマチンは間に立ってその両方をコントロールする。従って例えばOct4遺伝子のCD45を見ると緑の(H3K4me3)状態にあるものはなく、赤の(H3K27me3)状態にあるDNA断片がたくさんあることを示している。その下のSTAP細胞を見ると赤はなく緑がたくさんある。つまりOct4遺伝子のプロモーター領域が活性化されているので結果的にはOct4遺伝子を発現させてOct4蛋白質をたくさん産出しているということになる。この状態でOct4-GFPが組み込まれていると細胞は蛍光することになる。
ChIP seqに限らず、この遺伝子解析結果を重視する人々、Ts.Markerさん、DORAさん、学さん、和モガさん、そしてOoboeさんとそのパートナーさんが"STAP有る派"となっていることは衆知の事です。そして彼らは同時にキメラはナイフ切り分けであれ、スタンダードなやり方で論文通りに出来ていると考えている。若山さん本人が太田ESのコンタミだと言って明らかな嘘をついて論文を否定した理由は何らかの陰謀であると考えておられるらしいが、それがどういうものかに関して何ら推論も論証も無い。
対して小保方細胞核使用ntES論は説明した通りである。
1.キメラはntESでできている。
2.幹細胞もntESでできている。
3.STAP酸浴細胞は小保方さんが作った本物である。
丹羽論文はティシュー論文の一部を再現したが、理研での酸浴Oct4-GFP蛍光細胞が本物のOct4遺伝子をほんのわずかしか含まず、GFP蛍な光の漏れ出し現象を指摘した。それに対して、このLetter Figure 4-bの実験結果はたくさんの内在性Oct4遺伝子の発現を定量的に立証している。つまり小保方さんの再現実験があまりに過酷で十分な再現条件でなかった可能性を示している。
以下は彼女の再現実験の予備実験時の写真です。旧DORAブログにあるものですが、近々閉鎖になりますから、あちこちに証拠保存しておくという意味でも貼り付けて置きましょう。参考までに、後にOoboe さんのパートナーさんの取り寄せた、赤色フィルター切り替えで自家蛍光の無いことの確認されている、全てが、緑色スフィア塊の写真も西岡さんのガンバレブログに楠本さんが貼り付けています。
しかし、だからと言って、キメラができたことにはならない。
さて、一方で、レター論文にNBCI登録されていると書かれているのは以下の分ですが、上述したChIP seq分、或はここにも書かれているChIP-seq files はここに挙げられているリストにはない。
>>
RNA-seq and ChIP-seq files have been submitted to the NCBI BioSample databases under accessions SAMN02393426, SAMN02393427, SAMN02393428, SAMN02393429, SAMN02393430, SAMN02393431, SAMN02393432, SAMN02393433, SAMN02393434 and SAMN02393435.このリストに対応している登録サンプルは以下です。
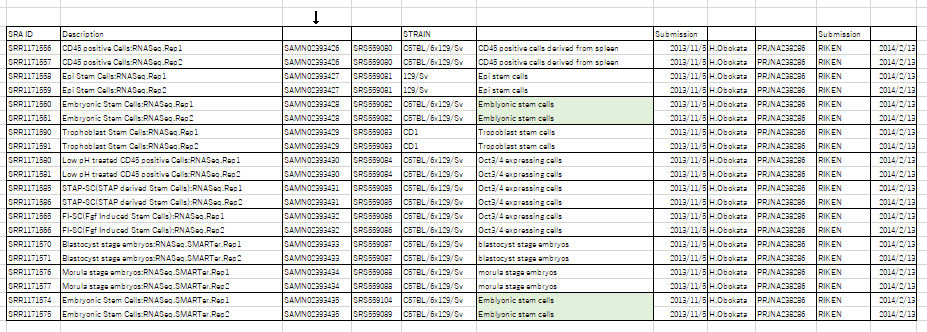
こちらのRNA分析にはES細胞が二つ登録されています。恐らく若山さんの作った分と、レター論文で使われたBFPのES細胞ではないかと思われます。これは丹羽さんが作っているのではないか。何にも確認されていません。ほんとに杜撰な調査です。すべて小保方さんや丹羽さんに聞いたら分かる事ばかりです。
報告書は二年前の2012年8月にGRASに提出されたLetter Figure 4-bのSTAPのグラフのもととなった上記⑫の生資料が保存されていたと説明しました。桂報告書16Pです。
>>
3)STAP 細胞由来 ChIP-seq (input)サンプルは 129B6 F1ES1 から取得された
(調査結果) 小保方氏が CDB ゲノム資源解析ユニット(以下「GRAS」という)に持参し残されてい た STAP 細胞由来 ChIP-seq (input)サンプルを再度 NGS 解析した結果、STAP 細胞由来と される ChIP-seq input データは CAG-GFP の挿入を持つ 129xB6 へテロ系統由来の細胞か ら取得されたものと判明した。さらに SNPs の解析、特異的な欠失変異の解析により(2 -3-1-1(d)参照のこと)CAG-GFP が挿入された 129B6 F1ES1 とほぼ同一細胞由来の データであることが明らかとなった。 ⑫があるにも関わらず、調査チームは保存されていたinputデータの混合液をもう一度解析したというのです。公共データ登録されているデータとどう違うかの説明はありません。inputデータは提出された細胞のDNA、RNAの全てを含んだ混合液のシーケンスデータです。その混合液自体が残されていて、これを他のゲノムと同量となる30倍になるまで読んだという伊藤さんの説明ですが、ど素人なのでその意味は分からない。
このSNPsの解析表がBCA報告の(Extended Data Fig. 2c)ですが、129B6F1 ES1が無いということは既に指摘しています。129B6F1 ES6とSTAP ChIP lysateはど素人が見ても全くパターンが違います。一方、AC129-1は129B6F1 ES1と同じだと言われているんですが、そのAC129-1とSTAP ChIP lysateとの比較はどうかというと、これはとても初期段階でTs.Marker さんが問題提起しました。
Ts.Marker さんは後にグラフィックスの精度の問題の範囲内かもしれないとおっしゃってますが、本日(2019/8/14)学ブログに以下の書き込みがありました。以前から指摘されていることですが、1Mbの範囲内で半分ほど違ってると改めて指摘されると、確かにこれは大きいですね。今のところSTAP ChIP lysate≠AC129-1(≒?129B6F1 ES1)ということではないでしょうか。
>>
拡張データ2CのSNPパターンの図ですが、縦の1枠(白抜きになっている部分)が1Mb(塩基配列100万個)です。縦枠150個並べて第6染色体の150Mbの全塩基を表しています。ChIP-seqとAC129は67Mb~68Mbの1Mbの範囲に違いがあり、AC129は全てB6ホモだけど、ChIP-seqは約500kbがB6ホモ、残りが129/B6ヘテロになっています。FLSの欠失は8番染色体で17kb、3番染色体の欠失は5kbで同じ欠失があることでFES1他と同一認定されましたが、違いが500Kbもあるというのは大きな違いです。細胞株が違うことを証明するのは、違いを1つ示せばいいので、これで細胞株が違うことは証明できています。
2019/8/14(水) 午前 9:56[ カツラ報告書 ]返信する黒線のところが1Mb幅で半分のSNPs数の違っているところです。
最初に指摘して置いたように、若山さんはややこしいことに取り下げ理由書にLetter Figure 4-bのSTAP細胞とES細胞のデータが取り違えられていると書いています。再掲します。
>>
(4) In Fig. 4b of the Letter, STAP cell and ES cell are wrongly labelled in a reverse manner.この取り違えは作表時なのか、サンプル提出時なのかわかりません。STAP ChIP lysateが提出時のES細胞の取り違えだったのなら、ここから129B6F1ESらしきものが出ても変ではありません。
ではその際は間違えられた方のESがSTAPだということになるが、そのマウス背景は何なのか。データ登録されたマウス背景はCD45がGOFで、TSがCD1、そしてその他は全部B6 X 129です。そしてそれを確かめるためのinputデータは揃っているが、検証はされていない。凍結lysateはSTAPだけだったのかどうかもわかってない。つまり、何事も系統立てては調べられていない。調査はとても恣意的で場当たり的だと言わざるを得ない。
- 2019/08/27(火) 09:34:47|
- AC129
-
-
| コメント:0
頑張れFBにパートナーのお知らせは取り敢えずのお知らせ内容でしたので、後日詳細をお伝えするとのことでしたが、誤解のおそれがありますので、少しだけ此方で但し書をします。
神戸地方検察庁には特別捜査部(特捜)と公安部の機能を兼ねた特別刑事部という部署があります。公安事案、労働事案、財政経済事案 等を扱う部署です。
今回、特別刑事部への情報申告事案は科学事案ではありません。
Stapの真偽についての情報申告ではなく社会的処理に於ける犯罪行為の存在疑義の情報申告であります。
その行為結果により、未来への可能性を示していたstap研究の芽を潰されてしまった事への真相究明の情報申告行為です。
2019/8/24(土) 午後 7:04[ Ooboe ]返信する
パートナーは、一週間前、申告内容を特別刑事部の担当官にご理解、把握して頂く為に前もって申告書と資料を送付していました。
申告書の標題は、検察による独自捜査の端緒を切っていただきたく犯罪疑義事案を申告いたします。
この申告書では刑事訴訟法第191条に基付いて特別刑事部の編成対応をもって捜査に取り組んで行かれることを申告いたします、と明記しています。
担当下さったのは、特別刑事部の窓口責任者の統括捜査官という肩書の方だったそうです。パートナーの大量の情報資料を読破されていたそうです。
ですので、パートナーの説明のご理解が速やかだったそうです。その結果、パートナー情報申告は十分な整合性の信憑性を有しているとのご判断をいただき実際に捜査する特検(特捜)に上げるということになったそうです。
また、この情報申告をネットやメディアに公表なさっても結構ですよ。と、意外な言葉にパートナーはビックリしてしまったそうです。
2019/8/24(土) 午後 7:53[ Ooboe ]返信する
**********
18. 小野小町 2019年08月26日 18:06
Ooboe さん、お元気?
パートナーさん、とうとうやり遂げたのね。検察がどう判断するかだわね。
学さんのところは引っ越し準備で書き込めないみたいね。ここのジムさんのところか根本さんのところを使わせて貰うといいわね。根本さんは法律の専門家のようよね。
一段落したらパートナーさんに早くブログ開設なさってとお願いしといてね。分かってる人を横に手伝ってもらったら早いわ。私たちは喜んでコメント欄に参加するから。
ところで差し支えなければちょっとお聞きしときたいんだけど、今回の特別刑事部への犯罪嫌疑申告というのは桂報告書に対するものと理解して置いていいのかしら。誰が嫌疑対象者なのかしら。右代表桂さんということかしら?
19. Ooboe 2019年08月27日 08:31
すみません、
嫌疑対象者氏の名義は、残念ながら
明示しないよう。との担当官のご注意だそうです。それと、著作権のもの以外、今回の資料の
申告含めUPは、ネット、メディアでもokだそうです。
20. Ooboe 2019年08月27日 09:08
あっ、はい、桂報告書に関わる嫌疑です。
**********
22. 小野小町 2019年08月27日 16:49
Ooboe さん、了解よ。応援しているわ。小保方さんが若山さんの目をかいくぐって既存ESを渡すことはできないわ。もう証明済みよ。ただ、犯罪が構成されているかどうか、そして諸般の事情を検察がどう判断するかしらね。私としては早稲田大学が小保方さんの博士号を卑劣な手段で奪ったことは許しがたいわ。学者の世界は料簡の狭い世界みたいだから日本での復帰は無理じゃないかと思うけど、博士号を持っていることは海外での研究活動には不可欠ね。桂報告書は無実の小保方さんから何もかも奪ってしまったわね。何かあったらおっしゃってね。力になれることがあれば協力するわ。じゃね。
23. Ooboe 2019年08月29日 01:58
小町さん
応援ありがとう😆💕✨
現在、特別刑事部の部長に上がっているとのことです。
約30ページの時系列説明書を精査されていると思います、
パートナーは頃合いを測って、それらの時系列根拠資料集を送付するの予定
パートナーは時効成立事案を含め時系列的に三次に渡る共謀画策に分類しており、その過程で複数発生していた犯罪行為につき、2件に絞り申告しました。
**********
コメント
おはようございます☀️
パートナーは、やっとのこと、
ワードにチャレンジ、
一般の方々が認識しやすい説明案内表示と
証拠文書をセットにして、楠本さんに
UPしてもらうそうです。が
作成苦戦中です、🤷🤦💨💢💦
2019/11/27(水) 08:39:46 Ooboe
パートナーは、小保方stap研究を無きものにした画策を第一次、第二次、第三次の画策として分類した時系列説明資料を、検察に提出しています。
この中での犯罪事案は諸々存在していましたが、偽計業務妨害、背任、公務員法違反、公文書虚偽作成、告発幇助間接正罪などのうち、明確なもの、2件にしぼって、告発手続きでなく、申告書提出の手続きをしているところです。
検察からの文書も全てではありませんが、一部公開する予定です。担当官は公開okとのことです。
2019/11/27(水) 13:30:12 Ooboe
第一次共謀画策は、2014年3月~6月16日
第二次共謀画策は、6月25日~12月26日
第三次共謀画策を、2015年1月~6月
と大きく分類しています。
この経緯の中で、分水嶺時点を6月5日として検察に資料を集中して説明しています。
そのパートナー証拠の更なる裏付けをお願いしていますが、頑張れFBでは、詳細は控えるようです。
全経緯を通じての、解明ポイントを6月5日の理研本部会議時点として捉えると、6月25日以後の第二次共謀画策や第三次共謀画策の経緯の本質が浮上してくるので、犯罪事案の、間接的ではあるが、背景として、その内容を検察に理解して貰うのに焦点をあてたとのことです。
この詳細は、ブログ開設で展開したいそうですが、検察には、この首謀者と共謀者を証拠をもって特定して提示しているとのことです。
この首謀者については、居士さんの考察とは異なるものとなっています。
第一次画策以前の2011年から2015年の第三次画策の全経緯を通じて一貫した首謀者としてパートナーは検察に提示して説明しています。
抽象的で、ごめんなさい。
2019/11/28(木) 00:33:12 Ooboe
*****
現在、FES1、とFES2の調査用のES細胞サンプルの考察が続いておられますが、パートナーが入手しているそのサンプルについての情報をお伝え致します。参考にして下さいませ。
2019/12/04 URL 編集
Ooboe
前コメント、名前入力わすれました。ごめんなさい。
FES1などについてのパートナー入手情報
理研広報は、調査サンプルを受領し、解析を担当した、CDB非対称細胞分裂研の研究者に直々に確認され、その研究者はFES1とntESG1は山梨大、若山教授から取り寄せました。
FES2とntESG2は山梨、若山教授とは別のところから取り寄せました。
これと同じ内容は、情報公開室が確認しています。
今晩、続きをお伝え致します。
2019/12/04 URL 編集
Ooboe
続きです。
前記にありますように、理研広報や調査委員会事務局に調査用サンプルの取り寄せについて報告された、BCA論文の筆頭著者でもあった研究者は、BCA論文中では、大田氏からFES1、FES2、ntESG1、ntESG2、を取り寄せたことになる記述をしています。
世界に発信されたBCA論文と、食い違う内容報告を、理研広報など、理研組織内では共有確認しているわけですが、なにより取り寄せ依頼し、受領し、解析したご当人がこの食い違いについての諸々の事実を把握していると思われます。
2019/12/04 URL 編集
Ooboe
さて、調査用ESサンプルを取り寄せ依頼、受領、解析をされた、CDB研究者はなにゆえに、理研にとっての公式発表を担当する広報室や、法令遵守担当のコンプライアンス室などの対し、組織内部にだけに共有される情報を伝え、対外発表のBCA論文や、調査報告書とは食い違う報告をしたのでしょうか?
もし、広報などに伝えた、内容通りならFES1とntESG1は、山梨大で用意された調査用サンプルということになります。
そして、FES2と、ntESG2は、別のところということですから。FES1とFES2は、出所が違う訳であり、細胞作成された、場所も同じでない別々に作られた、ことになります。又は、株分けでしょうか?
いずれにしろ、FES1は、京都大、大田氏から、取り寄せたのではなく、山梨大、若山研から取り寄せたと、BCA筆頭著者は、理研内部組織に伝えていたこの事実は、客観的公的事実であることには、変りがございません。
2019/12/05 URL 編集
Ooboe
パートナーは、FES1などのサンプル受領についての、MTA契約文書の開示請求をしましたが、情報公開室は、解析担当研に確認した結果を、パートナー宛に回答書を送付しています。その記述の中に、解析担当研究者が若山教授に直接依頼したもので、MTAや取り寄せ手続きの公的文書等は交わしていません、とありました。
6月30日理研は機関として、コンプライアンス室が予備調査を開始するとの発表をしていますが、しかしFES1等の調査取り寄せに予備調査事務局は、関与していませんでした。
この事案は普通の研究者間でのサンプル提供、受領手続きではありません。機関として、公的に取り寄せるべき事案であった訳ですから、関与していれば公的事務局として、文書交付は為されたはずでした。
解析調査をする側は、サンプルを取り寄せの依頼するのですから、取り寄せ宅配経費は、依頼側が支払うはずです。しかし解析担当研は会計処理をされてませんし、又、山梨大学、若山研の会計処理も為されてませんので、若山氏が自費でFES1の宅配経費負担をして理研解析担当研究者に送付していたことになります。
ということから、依頼したのは、逆に若山氏側であった。というのが整合的に帰結されてくる訳です。
以上
FES1等については不明瞭な事実が更に重なって存在していますが、省きます。
いずれにいたしましても、真正な調査サンプルとしての資格が無いに等しいのが、 FES1等サンプルでありました。
2019/12/05 URL 編集
**********
- 2019/08/24(土) 23:14:22|
- Ooboeさん情報
-
-
| コメント:0
博論第3章参照
(原文)
3.6 References
1. Berrier, A. L.; Yamada, K. M. Cell-matrix adhesion. J. Cell. Physiol.213:565□573: 2007.
2. Braam, S. R.; Zeinstra, L.i Litjens, S.; Ward-van Oostwaard, D.; van den Brink, S.; van Laake, L.; Lebrin, F.; Kats, P.; Hochstenbach, R.; Passier, R.; Sonnenberg, A.; Mummery, C. L. Recombinant vitronectin is a functionally defined substrate that supports human embryonic stem cell self-renewal via alphavbetaS integrin. Stem Cells 26-2257□2265: 2008.
3. Brakebusch, C.; Fassler, R. The integrin-actin connection, an eternal love affair. EMBO J. 22:2324□2333: 2003.
4. Busser, B. W.; Bulyk, M. L.; Michelson A. M. Toward a systems-level understanding of developmental regulatory networks. Curr. Opin. Genet. Dev. 18:521□529: 2008.
5. Casella, J. F.; Flanagan, M. D.; Lin, S. Cytochalasin D inhibits actin polymerization and induces depolymerization of actin filaments formed during platelet shape change. Nature 293-302□305: 1981.
6. Ceradini, D. J.; Kulkarni, A. R.l Callaghan, M. J.; Tepper, O. M.I Bastidas, N.;Kleinman, M. E.; Capla, J. M.; Galiano, R. D.; Levine, J. P.I Gurtner, G. C. Progenitor cell trafficking is regulated by hypoxic gradients through HIF-1 induction of SDF-1. Nat. Med. 10-858□864: 2004.
7. Chelberg, M. K.I Tsilibary, E. C.l Hauser, A. R.l McCarthy, J. B. Type IV coUagen-mediated melanoma cell adhesion and migration: involvement of multiple, distinct domains of the collagen molecule. Cancer Res. 49:4796□4802; 1989.
8. Chiou, S. H.; Kao, C. L.; Peng, C. H.; Chen, S. J.; Tarng, Y. W.; Ku, H. H.; Chen, Y. C.; Shyr, Y. M.; Liu, R. S.; Hsu, C. J.; Yang, D. M.; Hsul W. M.; Kuo, C. D.; Lee, C. H. A novel in vitro retinal differentiation model by co'culturing adult human bone marrow stem cells with retinal pigmented epithelium cells. Biochem. Biophys. Res. Commun. 326:578□585; 2005.
9. Choi, J. S.; Yang, H. J.; Kim, B. S.; Kim, J. D.; Kim, J. Y.; Yoo, B.; Park, K.; Lee, H. Y.; Cho, Y. W. Human extracellular matrix (ECM) powders for injectable cell delivery and adipose tissue engineering. J. Control. Release 139:2□7; 2009.
10. Cooper, H. M.; Tamura, R. N.; Quaranta, V. The major laminin receptor of mouse embryonic stem cells is a novel isoform of the alpha 6 beta 1 integrin. J. CellBiol. 115:843□850; 1991.
11. Czirok, A.; Zamir, E. A.; Filla, M. B.; Little, C. D.; Rongisli, B. J. Extracellular matrix macroassembly dynamics in early vertebrate embryos. Curr. Top. Dev. Biol. 73:237□258; 2006.
12. Decline, F.; Rousselle, P. Keratinocyte migration requires alpha2beta1 integrin-mediated interaction with the laminin 5 gamma2 chain. J. Cell Sci. 114:811□823; 2001.
13. Desban, N.; Lissitzky, J. C.; Rousselle, P.; Duband, J. L. alpha1beta1-integrin engagement to distinct laminin-1 domains orchestrates spreading, migration and survival of neural crest cells through independent signaling pathways. J. Cell Sci. 119:3206□3218; 2006.
14. Goto, M.; Sumiyoshi, H.; Sakai, T.J Fassler, R.! Ohashi, S.j Adachi, E.. Yoshioka, H.; Fujiwara, S. Ehmination ofepiplakin by gene targeting results in acceleration ofkeratinocyte migration in mice. Mol. Cell. Biol. 26:548□558; 2006
15. Hayashi, Y.; Furue, M. K; Okamoto, T.: Ohnuma, K.; Myoishi, Y.: Fukuhara, Y.: Abe, T.; Sato, J. D.; Hata, R.; Asasliima, M. Integrins regulate mouse embryonic stem cell self-renewal. Stem Cells 25-3005□3015; 2007.
16. Hehlgans, S.; Haase, M.; Cordes, N. Signalling via integrins: implications for cell survival and anticancer strategies. Biochim. Biophys. Acta 1775:163□180; 2007
17. Hynes, R. O. Integrins: bidirectional, allosteric signaling machines. Cell 110:673□687; 2002.
18. Khoshnoodi, J.; Pedchenko, V.; Hudson, B. G. Mammalian collagen IV. Microsc. Res. Tech. 71:357□370; 2008.
19. Kim, Y. S.; Park, H. J.; Hong, M. H.; Kang, P. M.; Morgan, J. P.; Jeong, M. H.; Cho, J. G.: Park, J. C.; Ahn, Y. TNF-alpha enhances engraftment of mesenchymal stem cells into infarcted myocardium. Front. Biosci. 14:2845□2856: 2009.
20. Kjtaori, T.; Ito, H.; Schwarz, E. M.: Tsutsumi, R.; Yoshitomi, H.; Oishi, S.;Nakano, M.; Fujii, N.; Nagasawa, T.: Nakamura, T. Stromal cell-derived factor 1/CXCR4 signaling is critical for the recruitment of mesenchymal stem cells to the fracture site during skeletal repair in a mouse model. Arthritis Rheum. 60:813□823; 2009.
21. Kofidis, T.; de Bruin, J. L.; Yamane, T.: Balsam, L. B.; Lebl, D. R.; Swijnenburg, R. J. :Tanaka, M.; Weissman, I. L.; Robbins, R. C. Insulin-like growth factor promotes engraftment, differentiation, and functional improvement after transfer of embryonic stem cells for myocardial restoration. Stem Cells 22;1239□1245; 2004.
22. Kofidis, T.: de Bruin, J. L.: Yamane, T.; Tanaka, M.; Lebl, D. R.; Swijnenburg, R. J.; Weissman, L L.; Robbins, R. C. Stimulation of paracrine pathways with growth factors enhances embryonic stem cell engraftment and host-specific differentiation in the heart after ischemic myocardial injury. Circulation 111:2486□2493: 2005.
23. Laflamme, M. A.: Chen, K. Y.; Naumova, A. V.; Muskheli, V.; Fugate, J. A.; Dupras, S. K.; Reinecke, H.; Xu, C.; Hassanipour, M.; Pohce, S.; O'Sullivan, C.; Collins, L.; Chen, Y.; Minami, E.; Gill, E. A.; Ueno, S.; Yuan, C.; Gold, J.; Murry, C. E. Cardiomyocytes derived from human embryonic stem cells in pro-survival factors enhance function of infarcted rat hearts. Nat. Biotechnol. 25:1015□1024: 2007.
24. Lee, J. M.; Dedhar, S.; Kalluri, R.I Thompson, E. W. The epithelial-mesenchymal transition- new insights in signaling, development, and disease. J. Cell Biol. 172:973□981; 2006.
25. Lee, K. H.; Chuang, C. K; Wang, H. W.: Stone, L.; Chen, C. H.; Tu, C. F. An alternative simple method for mass production of chimeric embryos by coculturing deヌded embryos and embryonic stem cells in Eppendorf vials. Theriogenology 67;228□237; 2007.
26. Leitinger, B.; Hohenester, E. Mammalian collagen receptors. Matrix Biol. 26:146-155; 2007.
27. Li, K.; Chuen, C. K.; Lee, S. M.; Law, P.; Fok, T. F.; Ng, P. C.; Li, C. K; Wong, D.: Merzouk, A.; Salad, H.; Gu, G. J.; Yuen, P. M. Small peptide アnalogue of SDF-lalpha supports survival of cord blood CD34+ cells in synergy with other cytokines and enhances their ex vivo expansion and engraftment into nonobese diabetic/severe combined immunodeficient mice. Stem Cells 24:55□64: 2006.
28. Li, S.; Tanaka, H.; Wang, H. H.; Yoshiyama, S.; Kumagai, H.; Nakamura, A.; Brown, D. L.; Thatcher, S. E..;Wright, G. L.; Kohama, K. Intracellular signal transduction for migration and actin remodeling in vascular smooth muscle cells after sphingosylphosphorylcholine stimulation. Am. J. Physiol. Heart Circ. Physiol. 291:H1262□1272; 2006.
29. Lock, J. G.; Wehrle-Haller, B.i Stromblad, S. Cell-matrix adhesion complexes: master control machinery of cell migration. Semin. Cancer Biol. 18:65□76; 2008
30. Matsuda, R.: Yoshikawa, M.; Kimura, H.; Ouji, Y.;Nakase, H.; Nishimura, F.; Nonaka, J.; Toriumi, H.; Yamada, S.; Nishiofuku, M.; Moriya, K.; Ishizaka, S.; Nakamura, M.; Sakaki, T. Cotransplantation of mouse embryonic stem cells and bone marrow stromal cells following spinal cord injury suppresses tumor development. Cell Transplant. 18:39□54; 2009.
31. Mooney, D. J.; Vandenburgh, H. Cell delivery mechanisms for tissue repair. Cell Stem Cell 2:205□213; 2008.
32. Muny, C. E.; Keller, G. Differentiation of embryonic stem cells to clinically relevant populations: lessons from embryonic development. Cell 132:661□680; 2008
33. Nishikawa, S.; Jakt, L. M.; Era, T. Embryonic stem-cell culture as a tool for developmental cell biology. Nat. Rev. Mol. Cell Biol. 8-502□507; 2007.
34. Pan, G.; Thomson, J. A. Nanog and transcriptional networks in embryonic stem cell pluripotency. Cell Res. 17:42□49: 2007.
35. Parameswaran, K.; Radford, K.; Zuo, J.; Janssen, L. J.; O'Byrne, P. M.; Cox, P. G. Extracellular matrix regulates human airway smooth muscle cell migration. Eur. Respir. J. 24;545□55l; 2004.
36. Passier, R.; van Laake, L. W.; Mummery, C. L. Stem-cell-based therapy and lessons from the heart. Nature 453:322□329; 2008.
37. Perris, R.; Syfrig, J.; Paulsson, M.; Bronner-Fraser, M. Molecular mechanisms of neural crest cell attachment and migration on types 1 and IV collagen. J. Cell Sci. 106(Pt4):1357□1368; 1993.
38. Poschl, E.; Schlotzer-Schrehardt, U.; Brachvogel, B.; Saito, K.; Ninomiya, Y.: Mayer, U. Collagen IV is essential for basement membrane stability but dispensable for initiation of its assembly during early development.
(和訳)
3.6参照 <訳注:11jigen氏は本文中にリファランスナンバーが打たれていないと指摘しているが、それが編集中の草稿であることの何よりの証拠。>
1. Berrier, A. L.; Yamada, K. M. 細胞 - マトリックス接着。 Cell. Physiol.213:565□573: 2007.
2. Braam, S. R.; Zeinstra, L.i Litjens, S.; Ward-van Oostwaard, D.; van den Brink, S.; van Laake, L.; Lebrin, F.; Kats, P.; Hochstenbach, R.; Passier, R.; Sonnenberg, A.; Mummery, C. L. 組換えビトロネクチンは、機能的に定義された基質であり、アルファベータSインテグリンを介したヒト胚性幹細胞自己再生を支持する。 Stem Cells 26-2257□2265: 2008.
3. Brakebusch, C.; Fassler, R. インテグリンアクチンとのつながり、永遠の恋愛。EMBO J. 22:2324□2333: 2003.
4. Busser, B. W.; Bulyk, M. L.; Michelson A. M. 発達規制ネットワークのシステムレベルの理解に向けて。 Curr. Opin. Genet. Dev. 18:521□529: 2008.
5. Casella, J. F.; Flanagan, M. D.; Lin, S. サイトカラシンDはアクチン重合を阻害し、血小板形状変化の間に形成されるアクチンフィラメントの解重合を誘導する。 Nature 293-302□305: 1981.
6. Ceradini, D. J.l Kulkarni, A. R.l Callaghan, M. J.I Tepper, O. M.I Bastidas, N.;Kleinman, M. E.; Capla, J. M.I Galiano, R. D.I Levine, J. P.I Gurtner, G. C. 前駆細胞の輸送は、SDF-1のHIF-1誘導を介して、低酸素勾配によって調節される。 Nat. Med. 10-858□864: 2004.
7. Chelberg, M. K.I Tsilibary, E. C.l Hauser, A. R.l McCarthy, J. B. IV型コラーゲン媒介黒色腫細胞の接着および遊走:コラーゲン分子の複数別個領域の関与。 Cancer Res. 49:4796□4802; 1989.
8. Chiou, S. H.; Kao, C. L.; Peng, C. H.; Chen, S. J.; Tarng, Y. W.; Ku, H. H.; Chen, Y. C.; Shyr, Y. M.; Liu, R. S.; Hsu, C. J.; Yang, D. M.; Hsul W. M.; Kuo, C. D.; Lee, C. H. 成人ヒト骨髄幹細胞を網膜色素上皮細胞と共培養することによる新規試験管内網膜分化モデル Biochem. Biophys. Res. Commun. 326:578□585; 2005.
9. Choi, J. S.; Yang, H. J.; Kim, B. S.; Kim, J. D.; Kim, J. Y.; Yoo, B.; Park, K.; Lee, H. Y.; Cho, Y. W. 注射による細胞送達および脂肪組織工学のためのヒト細胞外マトリックス(ECM)粉末。 J. Control. Release 139:2□7; 2009.
10. Cooper, H. M.; Tamura, R. N.; Quaranta, V. マウス胚性幹細胞の主要なラミニン受容体は、α6β1インテグリンの新規アイソフォームである。 J. CellBiol. 115:843□850; 1991.
11. Czirok, A.; Zamir, E. A.; Filla, M. B.; Little, C. D.; Rongisli, B. J. 初期の脊椎動物の胚における細胞外マトリックスのマクロダイナミクス Curr. Top. Dev. Biol. 73:237□258; 2006.
12. Decline, F.; Rousselle, P. ケラチノサイトの移動は、ラミニン5γ2鎖とのα2β1インテグリン媒介相互作用を必要とする。 J. Cell Sci. 114:811□823; 2001.
13. Desban, N.; Lissitzky, J. C.; Rousselle, P.; Duband, J. L. 異なるラミニン-1ドメインへのα1β1インテグリンの関与は、独立したシグナル伝達経路を介した神経堤細胞の広がり、遊走および生存を調整する。 J. Cell Sci. 119:3206□3218; 2006.
14. Goto, M.; Sumiyoshi, H.; Sakai, T.J Fassler, R.! Ohashi, S.j Adachi, E.. Yoshioka, H.; Fujiwara, S. 遺伝子ターゲティングによるエプラパキンの絶滅により、マウスにおけるケラチノサイトの移動が加速される。 Mol. Cell. Biol. 26:548□558; 2006
15. Hayashi, Y.; Furue, M. K; Okamoto, T.: Ohnuma, K.; Myoishi, Y.: Fukuhara, Y.: Abe, T.; Sato, J. D.; Hata, R.; Asasliima, M. インテグリンは、マウス胚性幹細胞の自己再生を調節する。 Stem Cells 25-3005□3015; 2007.
16. Hehlgans, S.; Haase, M.; Cordes, N. インテグリンを介したシグナル伝達:細胞生存と抗癌戦略への影響。 Biochim. Biophys. Acta 1775:163□180; 2007
17. Hynes, R. O. インテグリン:双方向のアロステリックシグナルマシン。 Cell 110:673□687; 2002.
18. Khoshnoodi, J.; Pedchenko, V.; Hudson, B. G. 哺乳動物コラーゲンIV。 Microsc. Res. Tech. 71:357□370; 2008.
19. Kim, Y. S.; Park, H. J.; Hong, M. H.; Kang, P. M.; Morgan, J. P.; Jeong, M. H.; Cho, J. G.: Park, J. C.; Ahn, Y.TNF-アルファは、間葉系幹細胞の梗塞心筋への移植を促進する。 Front. Biosci. 14:2845□2856: 2009.
20. Kjtaori, T.; Ito, H.; Schwarz, E. M.: Tsutsumi, R.; Yoshitomi, H.; Oishi, S.;Nakano, M.; Fujii, N.; Nagasawa, T.: Nakamura, T. 間質細胞由来因子1 / CXCR4シグナル伝達は、マウスモデルにおける骨格修復の間に骨折部位への間葉系幹細胞の動員にとって重要である。 Arthritis Rheum. 60:813□823; 2009.
21. Kofidis, T.; de Bruin, J. L.; Yamane, T.: Balsam, L. B.; Lebl, D. R.; Swijnenburg, R. J. :Tanaka, M.; Weissman, I. L.; Robbins, R. C. インスリン様増殖因子は、心筋再生のための胚性幹細胞移植後の生着、分化および機能改善を促進する。 Stem Cells 22;1239□1245; 2004.
22. Kofidis, T.: de Bruin, J. L.: Yamane, T.; Tanaka, M.; Lebl, D. R.; Swijnenburg, R. J.; Weissman, L L.; Robbins, R. C. 成長因子によるパラクリン経路の刺激は、虚血性心筋損傷後の心臓における胚性幹細胞の生着および宿主特異的分化を増強する。 Circulation 111:2486□2493: 2005.
23. Laflamme, M. A.: Chen, K. Y.; Naumova, A. V.; Muskheli, V.; Fugate, J. A.; Dupras, S. K.; Reinecke, H.; Xu, C.; Hassanipour, M.; Pohce, S.; O'Sullivan, C.; Collins, L.; Chen, Y.; Minami, E.; Gill, E. A.; Ueno, S.; Yuan, C.; Gold, J.; Murry, C. E. 生存促進因子におけるヒト胚性幹細胞由来の心筋細胞は、梗塞したラット心臓の機能を高める。Nat. Biotechnol. 25:1015□1024: 2007.
24. Lee, J. M.; Dedhar, S.; Kalluri, R.I Thompson, E. W. 上皮間葉移行 - シグナル伝達、発生および疾患における新しい洞察。 J. Cell Biol. 172:973□981; 2006.
25. Lee, K. H.; Chuang, C. K; Wang, H. W.: Stone, L.; Chen, C. H.; Tu, C. F. 裸胚と胚性幹細胞をエッペンドルフバイアルに共培養することにより、キメラ胚の大量生産のための簡単な代替方法。Theriogenology 67;228□237; 2007.
26. Leitinger, B.; Hohenester, E. 哺乳類のコラーゲン受容体。 26:146-155; 2007.
27. Li, K.; Chuen, C. K.; Lee, S. M.; Law, P.; Fok, T. F.; Ng, P. C.; Li, C. K; Wong, D.: Merzouk, A.; Salad, H.; Gu, G. J.; Yuen, P. M. SDF-1αの小ペプチド類似体は、他のサイトカインとの相乗作用で臍帯血CD34 陽性細胞の生存を助け、それらの体外での増殖および非肥満糖尿病/重症複合免疫不全マウスへの移植を補強する。 Stem Cells 24:55□64: 2006.
28. Li, S.; Tanaka, H.; Wang, H. H.; Yoshiyama, S.; Kumagai, H.; Nakamura, A.; Brown, D. L.; Thatcher, S. E..;Wright, G. L.; Kohama, K. スフィンゴシルホスホリルコリン刺激後の血管平滑筋細胞における遊走およびアクチン再構築のための細胞内シグナル伝達。 Am. J. Physiol. Heart Circ. Physiol. 291:H1262□1272; 2006.
29. Lock, J. G.; Wehrle-Haller, B.i Stromblad, S. 細胞 - マトリックス接着複合体:細胞移動のマスター制御機構。 Semin. Cancer Biol. 18:65□76; 2008
30. Matsuda, R.: Yoshikawa, M.; Kimura, H.; Ouji, Y.;Nakase, H.; Nishimura, F.; Nonaka, J.; Toriumi, H.; Yamada, S.; Nishiofuku, M.; Moriya, K.; Ishizaka, S.; Nakamura, M.; Sakaki, T. 脊髄損傷後のマウス胚性幹細胞および骨髄間質細胞の同時移植は、腫瘍発生を抑制する。 Cell Transplant. 18:39□54; 2009.
31. Mooney, D. J.; Vandenburgh, H. 組織修復のための細胞送達機構。Cell Stem Cell 2:205□213; 2008.
32. Muny, C. E.; Keller, G. 胚性幹細胞の臨床的に関連性の高い集団への分化:胚発生からの教訓。 Cell 132:661□680; 2008
33. Nishikawa, S.; Jakt, L. M.; Era, T. 発生細胞生物学のためのツールとしての胚性幹細胞培養。 Nat. Rev. Mol. Cell Biol. 8-502□507; 2007.
34. Pan, G.; Thomson, J. A. 胚性幹細胞多能性におけるNanogおよび転写ネットワーク。 Cell Res. 17:42□49: 2007.
35. Parameswaran, K.; Radford, K.; Zuo, J.; Janssen, L. J.; O'Byrne, P. M.; Cox, P. G. 細胞外マトリックスは、ヒト気道平滑筋細胞の移動を調節する。 Eur. Respir. J. 24;545□55l; 2004.
36. Passier, R.; van Laake, L. W.; Mummery, C. L. 幹細胞に基づく療法と心臓からの教訓。Nature 453:322□329; 2008.
37. Perris, R.; Syfrig, J.; Paulsson, M.; Bronner-Fraser, M. 1型および4型コラーゲンにおける神経堤細胞の接着および遊走の分子機構 J. Cell Sci. 106(Pt4):1357□1368; 1993.
38. Poschl, E.; Schlotzer-Schrehardt, U.; Brachvogel, B.; Saito, K.; Ninomiya, Y.: Mayer, U. コラーゲンIVは基底膜の安定性に必須であるが、初期発生時の組成開始には必ずしも必要でない。
- 2019/08/24(土) 22:55:48|
- 小保方さんの論文
-
-
| コメント:0
博論第3章
草稿目次との対比
(11jigen引用箇所)
3.小細胞の特性
3.1序論 - 48
3.1.1幹細胞の分化能
3.2実験 - 48
3.2.1試験管内での細胞の分化能 - 48
3.2.2生体内での分化能 - 48
3.3結果 - 49
3.3.1試験管内での細胞の分化能 - 50
3.3.2生体内での細胞の分化能 - 51
3.4第三章のまとめ - 51
3.5討論 - 51
3.6参照 - 58
(原文)
3. DIFFERENTIATION POTENTIAL OF SMALL CELLS
3.1 Introduction
3.1.1 Differentiation potential of stem cells
One of definitions of stem cells is multidifferentiation potential. Also the degree of their stemness is determined by their differentiation potential. According to results of section 2, sphere forming cells expressed pluripotent cell markers. Therefore in this section, we aimed to confirm their differentiation potential in vivo and in vitro.
(和訳)
3. 小細胞の分化能<訳注:目次では3. Characterization of small cellsとなっているので、小保方さんがいろいろと編集中であることがわかる。これを見て草稿だと気づけない頭というのもどうなのか。>
3.1 序論
3.1.1 幹細胞の分化能
幹細胞の定義の一つは多分化能である。それらの幹細胞性の程度もまたそれらの分化能によって決定される。第2章の結果によると、球体形成細胞は多能性細胞マーカーを発現した。したがってこの章では、我々は生体内および試験管内での分化能を確認することを目的とした。
(原文)
3.2 Experimental<→Experiment>
3.2.1 In vitro Differentiation Assays.
In vitro differentiation assays were examined following the published differentiation culture conditions for murin ES cells.
Mesoderm lineage differentiation assay. Dissociated muscle cells were stained with anti-αmooth muscle actin antibody, anti-Myosin antibody and anti-Desmin antibody. Chondrocyte were stained with Safranin-0 and Fast Green. Osteocytes were stained with ALIZARIN RED S. After 21 days, adipocytes were stained with Oil Re 0.
Ectoderm lineage (Neural lineage) differentiation assay. Cells were plated on ortinin-coated chamber slides and incubated with anti-βIII Tubuin mouse monoclonal, anti-O4 mouse monoclonal antibody and anti-GFAP mouse monoclonal antibody.
Endoderm lineage (Hepatic) differentiation assay. Differentiated cells were detected by immunohistochemistory using anti-αfetoprotein mouse monoclonal antibody, anti-Albumin goat polyclonal antibody and anti-Cytokeratin 18 mouse monoclonal antibody. Results from immunohistochemistry were confirmed by RT-PCR.
(和訳)
3.2 実験
3.2.1 試験管内分化実験<訳注:目次では3.2.1 Differentiation potential of cells in vitroとなっているので、小保方さんがいろいろと編集中であることがわかる。これを見て草稿だと気づけない頭というのもどうなのか。>
試験管内分化検証は次の公表されているげっ歯類ES細胞用分化培養条件で調べられた。
中胚葉系統分化検査。解離された筋細胞は anti-αmooth muscle actin 抗体、anti-Myosin 抗体 及び anti-Desmin 抗体で染色された。軟骨細胞はSafranin-0および Fast Greenで染色された。骨細胞はALIZARIN RED Sで染色された。21日後に脂肪細胞をOil Re 0で染色した。
外胚葉系統(神経系統)分化検査。細胞はortininコーティングされたチャンバースライド上に播種され、anti-βIII Tubuin mouse monoclonal、anti-O4 mouse monoclonal antibody及びanti-GFAP mouse monoclonal antibodyで培養された。
内胚葉系統(肝)分化検査。分化した細胞はanti-αfetoprotein mouse monoclonal antibody、anti-Albumin goat polyclonal antibody及びanti-Cytokeratin 18 mouse monoclonal antibodyを使った免疫組織化学によって検出された。免疫組織化学の結果はRT-PCRによって確認された。
(原文)
3.2.2 In Vivo Differentiation.
Spheres were seeded onto biodegradable scaffolds and implanted into subcutaneous of NOD/SCID mice (Charles River laboratories). After 6 weeks, the implants were harvested and fixed with 10% formaldehyde, then examined by immunocytochemistry.
(和訳)
3.2.2 生体内分化<訳注:目次では3.2.2 Differentiation potential in vivoとなっているので、小保方さんがいろいろと編集中であることがわかる。これを見て草稿だと気づけない頭というのもどうなのか。>
スフィアは生分解性培地上に播種し、NOD / SCIDマウス<訳注:超免疫不全マウス> (Charles River laboratories社)の皮下に移植した。 6週間後、移植片を採取し、10%ホルムアルデヒドで固定した後に免疫細胞化学によって調べられた。
(原文)
3.3Results
3.3.1 Differentiation potential of cells in vitro
When representative bone marrow derived spheres were dissociated into single cells and exposed to three different differentiation media, the cells differentiated to express specific genes of the three lineages, Map2 (ectoderm), MyoD (mesoderm) and alpha-fetoprotein (AFP, endoderm) (Fig. 10).The addition of a neural differentiation medium to the in vitro environment of cells from bone marrow spheres, resulted in expression of pIII tubulin (a marker for neuron) (Fig. 11).
Alternatively, the addition of 20% fetal calf serum to the media resulted in the expression of markers representative of mesoderm; that is, a-smooth muscle actin (Fig. 11) as well as the mesenchymal cells, chondrocytes, osteocytes and adipocytes (Fig. 12). Thus, cells from spheres differentiated into all cell types of neural (neurons, oligodendrocytes and ghas) and mesenchymal stem cell lineage (chondrocytes, osteocytes and adipocytes). When exposed to a hepatocyte differentiation media the expression of a-fetoprotein (Fig. 11), was seen, suggestive of differentiation into endodermal tissue.
(和訳)
3.3 結果
3.3.1 試験管内での細胞の分化能
代表的な骨髄由来のスフィアを単一細胞に解離し、三つの異なる分化培地に移植したとき、細胞は、Map2(外胚葉)、MyoD(中胚葉)及びα-フェトプロテイン(AFP、内胚葉)の三系統の特定の遺伝子を発現して分化した(図10)。骨髄スフィアからの細胞の試験管内環境への神経分化溶剤の添加は、pIIIのチューブリン(ニューロンのマーカー)の発現をもたらした(図11)。
代わりに、媒体に20%のウシ胎児血清を添加すると、中胚葉を代表するマーカーの発現を生じた。即ち平滑筋アクチン(図11)更には間葉系細胞、軟骨細胞、骨細胞及び脂肪細胞である(図12)。このように、スフィアからの細胞は神経(ニューロン、オリゴデンドロサイトおよび膠細胞)と間葉系幹細胞系譜(軟骨細胞、骨細胞及び脂肪細胞)の全ての細胞型に分化した。肝細胞分化培地にさらすと、内胚葉組織への分化を示唆するaフェトプロテインの発現(図11)が見られた。
(原文)
3.3.2 Differentiation potential in vivo.
Bone marrow spheres and ES cells were transplanted subcutaneously into immune deficient mice to examine their tumor-initiating capacity. As a result, after 6 weeks ES cells formed a tumor. Spheres did not form tumor as big as ES cells did. We concluded that the proliferative potential of sphere cells was much weaker than that of ES cells (Fig. 13).
Next, we investigated if transplanted cells differentiated in vivo after transplantation. Transplanted cells were harvested after 6 weeks, and processed for immunohistochemical analyses. According to results of immunohistochemical analyses, spheres differentiated into tissues derived from three germ layers in vivo (Fig. 14).
(和訳)
3.3.2 生体内分化能<訳注:目次では3.3.2 Differentiation potential of cells in vivoとなっているので、小保方さんがいろいろと編集中であることがわかる。これを見て草稿だと気づけない頭というのもどうなのか。>
骨髄のスフィア及びES細胞が、それらの腫瘍形成能力を調べるために、免疫欠損マウスに皮下移植された。その結果、6週間後にES細胞は腫瘍を形成した。我々はスフィア細胞の増殖能がES細胞よりもはるかに弱かったと結論付けた(図13)。
次に我々は移植された細胞が移植後に生体内で分化しているかどうかを調査した。移植された細胞は6週間後に回収され、免疫組織化学的分析に供された。免疫組織化学的分析の結果によると、スフィアは生体内で三胚葉に由来した組織に分化した(図14)。
(原文)
3.4 Summary of section 3
♦ Spheres differentiated into cells derived from all three germ layers in vivo and in vitro
3.5 Discussion
Spheres differentiated into cells derived from three germ layers in vitro. It is yet answered that it was either differentiation or trans-differentiation. Also it is hard to refer the difference from in vitro differentiation potential of mesenchymal stem cells.
However, at least sphere forming cells enabled to generate various mature cells. In addition, in vivo differentiation assay proved that sphere forming cells were indeed stem cells, but distinct from ES cells in proriferative potential. The relationship between proliferative potential and differentiation potential has yet been understood. Spheres in this study showed differentiation potential which fulfill the critereia for both mesenchymal stem cells and neural stem cells. We believe that the spheres studied contain precursor cells to both mesenchymal and neural stem cells lineages.
It is important to note that cells described above, were propagated as non-adherent spheres, and are not known to exist in vivo. The in vitro behavior of cells in the spheres is likely to be very different from cells that reside in vivo. How these stem cells harbor in adult body and how they exert their potential
The spheres generated seemed to be composed of heterogenous populations of cells,with some markers expressed in some spheres, and other markers expressed in different spheres generated from cells isolated from the same tissue, at the same time. We believe that these differences also may be a function of the environment in which the cells were maintained.
(和訳)
3.4 第三章のまとめ
♦ スフィアは生体内および試験管内で、全三胚葉由来細胞に分化した。
3.5 討論
スフィアは試験管内で三胚葉に由来する細胞に分化した。それは分化または分化転換<訳注:すでに分化した細胞が別の細胞種に転換する現象>のいずれかであると分かっている。また間葉系幹細胞の試験管内分化能との違いに言及することは困難である。
しかし、少なくともスフィア形成細胞はさまざまな成体細胞の生成を可能にした。加えて、生体内分化実験はスフィア形成細胞が実際に幹細胞であることを証明したが、増殖能力においてES細胞と峻別された。増殖能と分化能との関係はもう理解されてきている。この研究におけるスフィアは間葉系幹細胞と神経幹細胞の両方の基準を満たす分化能を示した。私たちは研究しているスフィアに間葉系と神経系の両方の幹細胞系統への前駆細胞が含まれていると信じている。
上述の細胞が非接着性スフィアとして知られ、かつ生体内に存在することが知られていないことに留意することが重要である。スフィア内細胞の試験管内での挙動は生体内に存在する細胞とは非常に異なる可能性が高い。如何にこれらの幹細胞は成体の中にとどまり、そして如何に彼らの潜在能力を伸ばすのか。<訳注:構文が不完全なところも編集中の文章であることを示唆している。>
生成されたスフィアは、細胞の不均一な集団で構成されているようだった。同じ組織から単離された細胞から生成されていながら、同時に、いくつかのスフィアにはいくつかのマーカー発現が伴い、別のスフィアには他のマーカー発現があった。(訳注:ティシュー論文に同様の趣旨の記述がある。Spheres seemed to contain heterogeneous populations of cells, with some markers expressed in some spheres, and other markers expressed in different spheres generated from cells isolated from the same tissue, at the same time. 小保方さんはティシュー論文を下地に博論を書こうとしていることがよくわかる。)我々は、これらの違いはその中で細胞が維持されている環境の関係かなと考えている。
(原文)
Figure 10 in differentiation of bone marrow spheres
After 6 weeks of culture, cells change their figurations into those of cells representative of three germ layers.
Figure 11 In vitro differentiation assay of cells from 3 germ layers.
Marrowspheres were dissociated and plated in each appropriate medium. Cells from spheres,differentiated into cells representative of the three germ layers. Neural cells (left), muscle cells (middle) cells
, hepatocytes (right). Neurons stained with plll tubuline (left),. Muscle cells stained with a-smooth muscle actin (middle). Hepatocytes were stained with a-fetoprotein (right).
(和訳)
図10 骨髄スフィアの分化
6週間培養後、細胞はその形状を三胚葉を代表する細胞のものに変化させる。
図11 三胚葉由来細胞の試験管内分化実験。
骨髄スフィアを解離させ、それぞれ適切な培地に蒔いた。 スフィア由来細胞は三胚葉を代表する細胞に分化する。 神経細胞(左)、筋細胞(中)細胞<訳注:これもティシュー論文のの最後の方に似た以下の文章があって、それをコピペ編集しているから同じエラーが見逃されている。現代っ子の文章作法が原因。Differentiation assay from myospheres(C) into neural cells(i,ii,and iii),muscle cells(iv,v,and vi)cells,and hepatocytes(vii,viii,and ix). > ,肝細胞(右)。ニューロンはplllチューブリンで染色された(左)。筋肉細胞は平滑筋アクチンで染色された(中央)。 肝細胞はα-フェトプロテインで染色された(右)。
(原文)
Figure 12 Mesenchymal lineage differentiation.
Dissociated spheres were plated into serum-containing medium and cultured for 14-21 days. Plated cells differentiated into mesenchymal lineage cells even plated cells were from spheres derived from endoderm or ectoderm tissues.
Marrow spheres differentiated into condrocytes (A), adipocytes (B) and osteocytes (C). Pnemospheres differentiated into condrocytes (D), adipocytes (E) and osteocytes (F). Spinalspheres differentiated into condrocytes (G) and adipocytes (H).
(和訳)
図12 間葉系統の分化。
解離したスフィアを血清含有培地に播種し、14~21日間培養した。 培養された細胞は、培養された細胞が内胚葉または外胚葉組織由来のスフィアからのものであってさえ、間葉系細胞に分化した。
骨髄スフィアは、コンドロサイト(A)、脂肪細胞(B)および骨細胞(C)に分化した。 胚スフィアはコンドロサイト(D)、脂肪細胞(E)および骨細胞(F)に分化した。 脊椎スフィアはコンドロサイト(G)と脂肪細胞(H)に分化した。<訳注:(I)が抜けている。ここでも草稿だと分かる。>
(原文)
Figure 13 Teratoma forming assay
10(to the power of 7)bone marrow cells and ES cells were injected subcutaneously into immunedificienl mice.
After 6 weeks of implantation, cell masses were harvested.
Figure 14 Teratoma like mass from bone marrow spheres contained nerve expressing betalll-tubuline (left)(ectoderm), muscle expressing desmin (middle)(mesoderm) and duct like structure expressing AFP (right)(endoderm).
(和訳)
図13 テラトーマ形成実験
10の7乗個の骨髄細胞およびES細胞を免疫不全マウスに皮下注射した。
移植6週間後、細胞塊を採取した。
図14 骨髄スフィア由来のテラトーマ様腫瘤は、ベータⅢチューブリンを発現する神経(左)(外胚葉)、デスミンを発現する筋肉(中)(中胚葉)およびAFPを発現する管様構造(右)(内胚葉)を含んでいた。
- 2019/08/24(土) 22:54:44|
- 小保方さんの論文
-
-
| コメント:0
博論第2章
草稿目次との対比
(11jigen引用箇所)
2.小細胞の単離
2.1序論 - 27
2.1.1幹細胞と小細胞 - 27
2.1.2幹細胞と球体形成 - 27
2.2実験 - 28
2.2.1小細胞の分離 - 28
2.2.2小細胞の特性 - 29
2.3結果 - 30
(11jigen非引用個所)
2.3.1単離細胞からの球体形成 - 30
2.3.2トリチュレーション効果 - 30
2.3.3小細胞の培養条件 - 31
2.3.4骨髄細胞からの球体形成 - 31
2.3.5未熟細胞マーカー発現 - 32
2.3.6胚的表現型の発現 - 33
2.3.7多能性マーカーの定量PCR - 34
2.4第二章のまとめ - 35
2.5討論 - 35
2.6参考文献 - 45
(原文)
2. ISOLATION OF SMALL CELLS
2.1 Introduction
2.1.1 Stem cells and small cells
Generally speaking, stem cells are small. The ratio of cytoplasm and nuclear is one of remarkable indexes of stem cells. However, the isolation method of only small cells is yet established.
(和訳)
2. 小細胞の単離
2.1 序論
2.1.1 幹細胞と小細胞
一般的に言えば、幹細胞は小さい。細胞質と核の割合は幹細胞の顕著な指標の一つである。しかし、小細胞だけの単離方法はもう確立されている。
(原文)
2.1.2 Stem cells and sphere formation
Sphere formation is recognized as one of the results of stem cell property since stem cell is recognized that they have strong proliferative potential and self-renewal potency. Recently, many reports demonstrated that various adult tissues contained sphere forming cells. Cells derived from many adult tissues, including retina, brain, cornea, olfactory neuroepithelium, pancreas, skin, muscle and bone marrow have been propagated as non-adherent clusters or spheres, as have embryonic stem cells.
Cells contained within the spheres described in those reports, exhibit neural lineage markers and appear to possess varying degrees of stem cell potency. We believe that adult stem cells described in various reports represent the same adult stem cells at different stages of development, expressing different degrees of potency. Sphere forming cells are much more immature than previously expected. We hypothesized that adult stem cells procured from any tissue, endoderm, mesoderm or ectoderm, could demonstrate a wide multipotency and cross germ layers when maintained in appropriate environments. Therefore, we first investigated how to isolate small cells. And then we examined if they had sphere forming potency in the serum-free condition.
(和訳)
2.1.2 幹細胞と球体形成
幹細胞が強い増殖能力および自己再生能力を持っていることが認識されて以来、スフィア形成は幹細胞特性の結果の一つとして認識されている。最近では多くの報告が、様々な成体組織がスフィア形成細胞を含むことを示している。網膜、脳、角膜、嗅覚神経上皮、膵臓、皮膚、筋肉および骨髄を含む多くの成人組織に由来する細胞が、胚性幹(ES)細胞がそうであるように、非付着性のクラスタ又はスフィアとして増殖されてきた。
これらのレポートに記載されているスフィアに含まれる細胞は神経系統マーカーを発現し、幹細胞能力の様々な程度を保有するように見える。我々は様々な報告書に記載された成体幹細胞は発達の異なる段階での効力の異なる程度を表現する同一の成体幹細胞を表すと考えている。スフィア形成細胞は前もって予想されていたよりもはるかに未成熟である。我々は内胚葉、中胚葉または外胚葉からのどんな組織から獲得された成体幹細胞でも、適切な環境下で維持するとき、広い多能性を示し、胚葉を横断することができるという仮説を立てた。そこで我々はまず小細胞を単離する方法を検討した。そして次に無血清状態でスフィア形成能があるかどうか調べた。
(原文)
2.2 Experimentall<→Experiment>
2.2.1 Isolation of small cells
Bone marrow, lung, muscle and spinal cord tissues were procured from 3- to 4-week-old C57BL/6J mice as described below. Bone marrow was acquired by flushing femur and tibia with culture media using an insulin syringe. Cells were plated at 1x10^ cells/cm" in F12/DMEM (1:1, v/v) supplemented with 2% B27, 20ng/ml bFGF and lOng/ml EGF
To isolate small cells from murin bone marrow, the following three types of methods were examined.
(和訳)
2.2 実験
2.2.1 小細胞の分離
以下に述べているように骨髄、肺、筋肉および脊髄組織は3~4週齢のC57BL / 6Jマウスから入手した。骨髄はインスリン注射器を用いて培養液で大腿骨と脛骨を洗い出すことによって取得した。細胞は1×10^細胞/ cmで、2%B27、20ng/ mlのbFGF及び10ng/ mlのEGFを補充したF12/ DMEM(1:1、v / v)に播種した。
げっ歯類骨髄の小細胞を単離するために、以下の3種類の方法が試された。(訳注:murin bone marrowは難波氏が噛みついてmiceという言葉も知らないのかと疑義されたが、無論murinはmiceより広い概念だというだけで間違いではない。キンガ論文にも使われている言葉。小保方さんは細胞シートの研究でよくこの言葉を使っていたので、こう書いて置いたかもしれない。何しろ何よりもこれは草稿であった。)
(原文)
A. Cell sorter
Forward scatter was calibrated by size-defined beads. Less than 8 micro meters in diameter cells were isolated.
B. Optimistic<→Osmotic> pressure
Bone marrow cells were exposed to low optimistic<→osmotic> pressure liquid to destroy mature cells.
C. Trituration using thin-glass pipette
Standard glass pipettes were burned and stretched out to make thin tips. Mature cells were passed through thin glass pipettes many times and destroyed by mechanical stress. Obtained small cells were cultured in serum free medium, and appeared spheres were counted as a number of stem cells.
(和訳)
A. 細胞ソーター
前方散乱はサイズ定義されたビーズにより調整された。直径8マイクロメートル未満の細胞が単離された。
B. 浸透圧<訳注:これも難波氏の噛みついたところ。単なる勘違いでお疲れというだけの話。無論osmoticの間違い。論文審査で直されるのでこれが草稿だという何よりの証拠の一つでもある。>
骨髄細胞は成熟細胞を破壊するために低浸透圧液体に曝された。
C. 細ガラスピペットを使用しての粉砕
標準ガラスピペットは細い先端を作るために焼かれて伸ばされた。成体細胞は細ガラスピペットを何度も通され機械的ストレスによって破壊された。得られた小細胞は、無血清培地で培養し、スフィアが幹細胞の数としてカウントされた。
(原文)
2.2.2 Characterization of small cells
Immunohistochemistry. Protein expression was assessed using immunohistochemical techniques as described below. Each slide was incubated with anti-c-kit rat monoclonal antibody, anti-Sca-1 rat monoclonal antibody or anti-E-cadherin rat monoclonal antibody. After washing with PBS, the cells were incubated with goat anti-rat IgG Texas Red-conjugated antibodies and goat anti-rat IgG Fluorescein-conjugated antibodies. SSEA-1 and Alkaline phosphatase (AP) staining was performed using the ES cell detection Kit
Single sphere RT-PCR. Single spheres were individually collected under the microscope. Total RNA was extracted from each single sphere, and then subjected to oligo-dT-primed reverse transcription (RT). RT-PCR was performed using TaqDNA polymerase on an iCycler for 35 cycles.
(和訳)
2.2.2 小細胞の特性
免疫組織化学。タンパク質発現は下記に記された免疫組織化学法を用いて評価された。各ガラススライドはanti-c-kit rat monoclonal 抗体, anti-Sca-1 rat monoclonal 抗体または、 anti-E-cadherin rat monoclonal 抗体によって培養された。 PBSで洗浄後、細胞は goat anti-rat IgG Texas Red-conjugated 抗体と goat anti-rat IgG Fluorescein-conjugated 抗体で培養された。ES細胞検出キットを用いてSSEA-1およびアルカリホスファターゼ(AP)染色が行なわれた。<訳注:ピリオドが無い。ここも草稿である証拠の一つ。そもそも小保方さんはかなり粗忽な性格で、ケアレスミスはとても多い。博論の草稿を印刷するというのはいくら多忙でも粗忽すぎる。文章も切り貼りして校正するときに動詞が抜けたりする。>
単体スフィアの逆転写ポリメラーゼ連鎖反応。単体スフィアが個別に顕微鏡下に集められた。全RNAが各単体スフィアから抽出され、次にオリゴdTプライミング逆転写(RT)が行なわれた。 RT-PCRはiCycler上のTaqDNAポリメラーゼを用いて35サイクル行われた。
(原文)
2.3 Results
2.3.1 Sphere forming from isolated cells
Trituration and optimistic<→osmotic> pressure produced spheres. Whereas, cell sorter did not produced any spheres. Trituration produced most number of spheres. Therefore, we utilized trituration as the isolation method of small cells.
(和訳)
2.3 結果
2.3.1単離された細胞からのスフィア形成
粉砕と浸透圧<ここでも続けてoptimisticとosmoticの取り違えがあるがoptimisticというのは楽観的という意味で誰でも知っている。こんな言葉とosmoticを取り違えたまま気づかないでいるということを疲れてるなと思わない人は悪意なだけの人。金をもらって悪口を書いている。>がスフィアを作った。一方、セルソーターはスフィアを作らなかった。<訳注:Whereasは副詞的に使用しているようだが前文とカンマとの関係がおかしい。草稿ということもあってこのような文章になっている。普通こういうところを読むと草稿だと気付くものだが難波氏や11jigenは悪意を持って読んでいるから気づかない。> 粉砕が最も多くスフィアを生産した。我々は粉砕法を小細胞の単離方法として採用した。
(原文)
2.3.2 Effect of trituration
After the trituration, the population of small cells was increased comparing to native bone marrow cells (Fig. 3A and B). However not all of cells over 8 micro meters in diameter disappeared. Triturated cells formed spheres during culture.Interestingly, spheres were consisted of only small cells (Fig. 3C). Thus, it was demonstrated that trituration enabled to culture and grow only small cells.
(和訳)
2.3.2 トリチュレーション効果
粉砕後、小細胞の集団は元の骨髄細胞と比較して増加した(図3AおよびB)。 しかし、直径8ミクロンを超える細胞のすべてが消失したわけではない。粉砕された細胞は培養中にスフィアを形成した。興味深いことに、球体は小さな細胞のみで構成されていた(図3C)。 したがって、粉砕は小細胞のみを培養増殖させ得たことが実証された。
- 2019/08/24(土) 22:53:49|
- 小保方さんの論文
-
-
| コメント:0
対訳博論目次
(原文)
Contents
1. Background
1.1 General Introduction - 1
• Importance of stem cells
• Unique properties of all stem cells
1.2 Adult stem cell - 7
• History of Adult stem cell research
• Function of adult stem cells
1.3 Research methods for identifying adult stem cells - 10
• Normal differentiation pathways of adult stem cells.
• Transdifferentiation.
1.4 Pluripotent stem cells - 14
• The similarities and differences between embryonic and adult stem cells
• Embryonic stem cells
A. Establish<→Estalishment> of embryonic stem cells grown in the laboratory
B. Tests for identifying embryonic stem cells
• Induced pluripotent stem cells
1.5 Possibility of existence of adult pluripotent stem cells - 20
• Spore-like stem cells
1.6 Hypothesis of this research - 23
2. Isolation of small cells
2.1 Introduction - 27
2.1.1 Stem cells and small cells - 27
2.1.2 Stem cells and sphere formation - 27
2.2 Experimental<→Experiment> -28
2.2.1 Isolation of small cells - 28
2.2.2 Characterization of small cells - 29
2.3 Results - 30
2.3.1 Sphere forming from isolated cells - 30
2.3.2 Effect of trituration - 30
2.3.3 Culture conditions of small cells - 31
2.3.4 Sphere formation from bone marrow cells - 31
2.3.5 Immature cell marker expression - 32
2.3.6 Expression of embryonic like phenotypes - 33
2.3.7 Quantitative PCR of Pluripotent markers - 34
2.4 Summary of section 2 - 35
2.5 Discussion - 35
2.6 References - 45
3. Characterization of small cells
3.1 Introduction - 48
3.1.1 Differentiation potential of stem cells
3.2 Experimental<→Experiment> - 48
3.2.1 Differentiation potential of cells in vitro - 48
3.2.2 Differentiation potential in vivo - 48
3.3 Results - 49
3.3.1 Differentiation potential of cells in vitro -50
3.3.2 Differentiation potential of cells in vivo - 51
3.4 Summary of section 3 - 51
3.5 Discussion - 51
3.6 Referances - 58
4. Isolation of small cells from TISSUES DERIVED FROM ALL THREE GERM LAYERS
4.1 Introduction - 63
4.2 Experimental<→Experiment> - 63
4.2.1 Isolation of small cells from various tissues - 64
4.2.2 Characterization of isolated small cells - 64
4.2.3 Differentiation potential of sphere forming small cells - 64
4.3 Results - 65
4.3.1 Sphere formation from tissues derived from representative of the three germ layers - 65
4.3.2 Immature gene expression of spheres derived from tissues representative of the three germ layers -66
4.3.3 Differentiation potential of Cells derived from tissues representative of the three germ layers-67
4.3.4 Differentiation potential of cells in vivo - 68
4.4 Summary of section 4 - 69
4.5 Discussion - 69
4.6 References - 76
5. Chimera mice generation from ISOLATED SMALL CELLS
5.1 Introduction - 81
5.1.1 Adult stem cells and chimera mice - 81
5.2 Experimental<→Experiment> - 81
5.2.1 Chimera mice generation with spheres - 82
A. Aggregation method
B. Injection method
5.2.2 Genotyping - 82
5.2.3 Immunohistological analyses - 82
5.3 Results - 83
5.3.1 Generation of chimera zygotes - 83
5.3.2 アnalyses of adult chimera mice - 83
5.3.3 アnalyses of chimera fetuses - 83
5.4 Summary of section 5 - 84
5.5 Discussion - 85
5.6 References - 93
6. Future prospects
6.1 General overview - 95
6.2 Cell source for tissue engineering regenerative medicine - 95
6.3 Future issues - 96
APPENDIX - 97
• Material and method
• Primer sequences
ACKNOWLEDGEMENTS
CURRICULUM VITAE
(和訳)
目次
1.背景
1.1概論 - 1
•細胞の重要性
•すべての幹細胞のユニークな特性
1.2成体幹細胞 - 7
•成体幹細胞研究の歴史
・成体幹細胞の機能
1.3成体幹細胞を同定するための研究方法 - 10
•成体幹細胞の正常な分化経路
•分化転換。
1.4多能性幹細胞 - 14
•胚性および成体幹細胞の類似点と相違点
•胚性幹細胞
A.実験室で増殖した胚性幹細胞の樹立
B.胚性幹細胞の同定試験
・誘導性多能性幹細胞
1.5多能性成体幹細胞存在の可能性 - 20
•胞子様幹細胞
1.6この研究の仮説 - 23
2.小細胞の単離
2.1序論 - 27
2.1.1幹細胞と小細胞 - 27
2.1.2幹細胞と球体形成 - 27
2.2実験 - 28
2.2.1小細胞の分離 - 28
2.2.2小細胞の特性 - 29
2.3結果 - 30
2.3.1単離細胞からの球体形成 - 30
2.3.2トリチュレーション効果 - 30
2.3.3小細胞の培養条件 - 31
2.3.4骨髄細胞からの球体形成 - 31
2.3.5未熟細胞マーカー発現 - 32
2.3.6胚的表現型の発現 - 33
2.3.7多能性マーカーの定量PCR - 34
2.4第二章のまとめ - 35
2.5討論 - 35
2.6参考文献 - 45
3.小細胞の特性
3.1序論 - 48
3.1.1幹細胞の分化能
3.2実験 - 48
3.2.1試験管内での細胞の分化能 - 48
3.2.2生体内での分化能 - 48
3.3結果 - 49
3.3.1試験管内での細胞の分化能 - 50
3.3.2生体内での細胞の分化能 - 51
3.4第三章のまとめ - 51
3.5討論 - 51
3.6参照 - 58
4.全三胚葉由来組織からの小細胞の単離
4.1序論 - 63
4.2実験 - 63
4.2.1様々な組織からの小細胞の分離 - 64
4.2.2分離された小細胞の特性 - 64
4.2.3球体を形成する小細胞の分化能 - 64
4.3結果 - 65
4.3.1三胚葉代表由来組織からの球体形成 - 65
4.3.2三胚葉代表組織由来球体の未熟遺伝子発現 - 66
4.3.3三胚葉代表組織由来細胞の分化能 - 67
4.3.4生体内における細胞の分化能 - 68
4.4第四章のまとめ - 69
4.5討論 - 69
4.6参考文献 - 76
5.分離小細胞からのキメラマウス作製
5.1序論 - 81
5.1.1成体幹細胞とキメラマウス - 81
5.2実験 - 81
5.2.1球体によるキメラマウス作製 - 82
A.凝集法
B.注入法
5.2.2遺伝子型決定 - 82
5.2.3免疫組織学分析 - 82
5.3結果 - 83
5.3.1キメラ接合体の作製 - 83
5.3.2成長キメラマウスの分析 - 83
5.3.3キメラ胎児の分析-83
5.4第五章のまとめ - 84
5.5討論 - 85
5.6参考文献 - 93
今後の展望
6.1概観 - 95
6.2組織工学的再生医療用の細胞供給源 - 95
6.3今後の課題 - 96
付録 - 97
•材料と方法
•プライマー配列
謝辞
履歴書
- 2019/08/24(土) 22:51:47|
- 小保方さんの論文
-
-
| コメント:0
対訳博論概要
早稲田大学大学院先進理工学研究科
博 士 論 文 概 要
論 文 題 目
Isolation of pluripotent adult stem cells
discovered from tissues derived from all three germ layers
三胚葉由来組織に共通した
万能性体性幹細胞の探索
申 請 者
Haruko
Obokata
小保方
晴子
生命医科学専攻 環境生命科学研究
2010年 12月
体性幹細胞は、成体の体内に実際に存在し、生体の恒常性を保つため老化細胞の代替となる若い細胞を生み出し、炎症などの生体反応に応答して失われた細胞を補充する役割を担っていると考えられている。現在までに、造血幹細胞、間葉系幹細胞、神経幹細胞は多種の分化可塑性を有する体性幹細胞として研究が進められている。また、前駆細胞との区別が難しいが、各種生体組織にはそれぞれの組織幹細胞が存在していると考えられており、多くは培養系においてその存在が認められている。間葉系幹細胞研究に代表されるように、体性幹細胞の研究は発生学的な観察に基づき展開されている。哺乳類の発生において三胚葉分化は決定的な過程であり、体性幹細胞の多くも三胚葉分化の後に存在が確認されることから、三胚葉分化が起こった後は、例えば外胚葉系組織に存在している幹細胞が中胚葉や内胚葉由来組織の細胞に分化する、中胚葉系に存在している細胞が外胚葉・内胚葉由来組織の細胞に分化すると言った、いわゆる胚葉を超えた分化は起こりえないと考えられている。しかし近年、分子生物学的解析手法の発展により間葉系幹細胞の一部は外胚葉系の細胞から構成されることや、間葉系幹細胞が生体内で神経形成に関与するなどといった、いわゆる胚葉を越えた分化が三胚葉形成の後にも起こっていることが報告されている。これらの報告により、体性幹細胞の起源や分化能の限界についての大前提に疑問がもたれるようになってきている。
Vacantiらは2000年に、全身の生体の組織内には三胚葉由来によらず非常に強いストレスに耐性を有するspore-like stem cellが存在し体性幹細胞の補充に寄与している可能性を提唱してきた。その後、他の研究グループからも同様な概念に基づいた研究報告が相次いでいる。2002年には骨髄中に万能性幹細胞MAPCが存在することが報告され、2004年には間葉系幹細胞の一部に分化万能性を有するMIAMI cellが存在することが報告され、2006年には造血幹細胞の小さいサイズの分画の中にVSELS cellsが存在することが報告され、2010年には間葉系細胞の一部にストレス耐性のmuse cellsが存在することが報告されている。本研究では、spore-like stem cellの仮説を証明する第一歩として、全身の組織に共通の性質を持つ幹細胞が存在することを証明することを目標とし、幹細胞の採取、解析、再生医療研究応用への可能性を検討した。第一章では、生体組織由来のpluripotent stem cellに関する研究の動向を概説し、本研究の背景をまとめると共に、本研究の意義及び目的を明らかにした。
第二章では、spore-like stem cellの採取法を検討すると共に、幹細胞マーカーの発現を解析した。Spore-like stem cellsは細胞直径が非常に小さいという特徴を有しているため、小さい細胞を採取する方法を施行した。まず、cell sorterを用いてBlack6 マウスの骨髄細胞から、直径6μm以下の細胞のみを回収した。続いて低浸透の溶液で細胞を短時間処理することによって、大きな細胞の細胞膜を破壊し小さな細胞のみを回収した。また先端を10μmほどまで細めたガラスピペットで細胞を粉砕することによって小さい細胞を回収した。それぞれの方法で回収した細胞群を無血清培地で培養を行うと浸透圧処理または粉砕処理によって回収された細胞群から浮遊した球形のコロニー形成(以降sphereと呼ぶ)が確認された。粉砕処理を行った場合、高頻度にsphere形成が観察された。sphere形成の数は年齢依存的であり、生後4週齢のマウスからは生後8週齢のマウスの約二倍の数が観察された。Sphere形成は幹細胞の強い自己複製増殖能の結果として現れる現象であると考えられているため、免疫染色により、幹細胞マーカーの発現解析を行った。まず間葉系幹細胞や造血幹細胞など広範な体性幹細胞に発現が報告されているc-kitとSca-1の発現を調べた結果、多くのsphereに発現が確認された。続いてES細胞や発生初期の受精卵に発現が観察される万能性幹細胞マーカーであるSSEA-1とE-cadherinの発現を調べた結果、これらも多くのsphereに発現が確認されることが明らかとなった。
タンパク質レベルでの万能性幹細胞マーカーの発現が確認されたことから、遺伝子レベルでの万能性幹細胞マーカーの発現をES細胞の遺伝子発現と比較して検討を行った。特に、タンパク質レベルでのマーカーの発現強度は個々のsphereによって異なっていたため、個々のsphereは異なる遺伝子発現パターンを示すのではないかと言う予想の下、sphere一つ一つを顕微鏡下で採取し遺伝子発現の解析を行った。その結果、Oct4, Nanog, Sox2, Ecat1, Cripto, Esg1など万能性幹細胞に特異的に発現が見られる遺伝子マーカーが高頻度に発現していることが分かった。
第三章では、これらの細胞の分化能をin vitro, in vivoの双方で調査した。ES細胞から三胚葉由来の細胞へ分化させるための培養条件を参考に、培養条件を設定し分化誘導実験を行った。その結果、sphere由来の細胞は神経・筋肉・肝実質細胞などの代表的な三胚葉由来組織細胞へ分化できることが確認された。生体内での分化能と増殖能を検討するために移植実験を行った。Sphereの細胞はPGA上に播種され、2-3日PGA(poliglycolic acid)上に細胞を接着させるために培養した後、NOD/SCIDマウスの皮下に移植した。4-6週間後に移植片を採取し、組織学的、免疫組織化学的に解析を行った。移植後直径3mmほどのカプセル化した塊を形成した。内部には上皮、神経、筋肉、管といった三胚葉由来すべての組織形成が確認された。以上の結果から、粉砕処理後にsphereを形成する細胞は、無血清条件下で培養すると、非常に幼弱なタンパク質・遺伝子を発現し、培養系、生体内双方において三胚葉系由来組織への分化能を有することが示された。
第四章では同様の細胞群がその他の組織にも存在しているかを確認するため三胚葉由来組織の代表的な組織である脊髄(外胚葉)、筋肉(中胚葉)、肺(内胚葉)から細胞を単離し、粉砕処理後、無血清培養条件下で浮遊培養を行った。タンパク質マーカーの発現は骨髄で行ったときと同様にc-kit, Sca-1, SSEA-1, E-cadherin陽性の細胞が確認された。遺伝子発現解析の結果、骨髄のときと同様、ES細胞に特異的な遺伝子の発現が多数確認された。特に肺由来のsphereからは高頻度にOct4陽性のsphere細胞塊が確認された。一方、脊髄からは多くのsphere形成が確認されるが、Oct4などのES細胞特異的な遺伝子マーカーを発現したsphereの割合は骨髄由来のsphereと比較して低い値を示した。培養系での分化誘導実験を行うと、骨髄のときと同様に、各特異的なマーカーで陽性を示す三胚葉由来組織の細胞へと分化した。さらにPGAに播種しNOD/SCIDマウスの皮下に移植すると、骨髄のときと同様に上皮、神経、筋肉、軟骨、腺といった三胚葉系の組織へと分化した。以上のことから、骨髄中から発見された広範な分化能を有する細胞群は、脊髄、筋肉、肺といったすべての三胚葉由来組織からも単離され得ることが確認された。
第五章では、幹細胞の万能性を証明するための最も重要な証明方法であるキメラマウスの作成を幼弱神経幹細胞培養条件であるbFGF, LIF依存浮遊培養系によって培養したsphereを用いて試みた。ICRマウスの受精卵とsphereを用いた凝集法によってキメラ卵を作成し、24時間培養した後、子宮に移植した。20日後に産まれた新生児の毛皮にはsphere由来の毛が観察されなかった。また産まれてきた新生児の数は移植した受精卵の数よりも少なかった。キメラの胎生致死、もしくは特定の組織への貢献、もしくは低頻度での貢献の可能性が考えられたため、胎生12.5日目の胎児の解析を行った。その結果全身にsphere由来の細胞が散在していることが確認された。このことから、sphere由来の細胞は全身の組織形成に寄与できる能力を有していることが明らかとなった。
第六章では本研究の総括と展望について述べた。本研究ではこれまでの常識を超えた体性幹細胞の起源と分化可能性についての新しい学説をもとに、体性幹細胞の創生の可能性を探る実験を試みた。本研究で得られた幹細胞が実際に生体内に存在するかどうかは、これから明確にすべき大きな課題である。しかしながら培養法をさらに効率化することによって大量培養を可能とし、組織工学をはじめとする再生医療研究の新たな細胞ソースとして期待できる。また、これまでiPS細胞を始めとした万能性幹細胞の創生の研究が盛んに行われているが、再生医療研究に必要なのは安全に機能する体性幹細胞であり、万能性幹細胞からの体性幹細胞の分化誘導は難しい。そこで本研究の第四章で検討したような体性幹細胞を体細胞から創出する試みが成功すれば細胞生物学的にも発生学的にも非常にインパクトのある研究成果となり、再生医療応用に最も適した細胞ソースを提供できるようになるものと期待される。
早稲田大学 博士(工学) 学位申請 研究業績書
氏 名 小保方 晴子 印
(2010年 11月 現在)
種 類 別
題名、 発表・発行掲載誌名、 発表・発行年月、 連名者(申請者含む)
(原文)
報文
(1) Haruko Obokata, Masayuki Yamato, Kohji Nishida, Satoshi Tsuneda, and Teruo Okano, "Subcutaneous transplantation of autologous oral mucosal epithelial cell sheets fabricated on temperature-responsive culture dishes", Journal of Biomedical Materials Research: A 86:1088-96. 2007
〇(2) Haruko Obokata, Koji Kojima, Masayuki Yamato, Satoshi Tsuneda, Charles A. Vacanti, “The potential of stem cells in adult tissues representative of the three germ layers.”,Tissue Eng Part A. 2010 in press.
(3) Haruko Obokata, Masayuki Yamato, Kohji Nishida, Satoshi Tsuneda, and Teruo Okano,“Time-course アnalyses of reconstructed ultrastructure change in oral mucosal epithelium cell sheet after subcutaneous transplantation” submitted to Journal of Tissue Engineering and Regenerative Medicine.
(4) Haruko Obokata, Masayuki Yamato, Satoshi Tsuneda, and Teruo Okano, “Protocol for reproducible subcutaneous transplantation of cell sheets” submitted to Nature Protocols.
(5) Haruko Obokata, Masayuki Yamato, Satoshi Tsuneda, and Teruo Okano, “The effectiveness of inflammatory cytokines for regeneration of transplanted cultured keratinocyte cells” submitted to Journal of Investigative Dermatology
(6) Haruko Obokata, Masayuki Yamato, Satoshi Tsuneda, and Teruo Okano, “Hyperacute inflammation アnalyses after subcutaneous transplantation of human fibroblast cell sheets into Lewis rats.”submitted to Xenotransplantation.
(和訳)
報文
(1) Haruko Obokata, Masayuki Yamato, Kohji Nishida, Satoshi Tsuneda, and Teruo Okano, 「温度応答性培養皿上で作製された自己口腔粘膜上皮細胞シートの皮下移植」, Journal of Biomedical Materials Research: A 86:1088-96. 2007
〇(2) Haruko Obokata, Koji Kojima, Masayuki Yamato, Satoshi Tsuneda, Charles A. Vacanti, 「三胚葉各成体組織内在幹細胞の潜在能力」,Tissue Eng Part A. 2010 in press.
(3) Haruko Obokata, Masayuki Yamato, Kohji Nishida, Satoshi Tsuneda, and Teruo Okano,「皮下移植後の口腔粘膜上皮細胞シートにおける再構築された超構造変化の時間経過分析」submitted to Journal of Tissue Engineering and Regenerative Medicine.
(4) Haruko Obokata, Masayuki Yamato, Satoshi Tsuneda, and Teruo Okano, 「細胞シートの再現性ある皮下移植のためのプロトコール」submitted to Nature Protocols.
(5) Haruko Obokata, Masayuki Yamato, Satoshi Tsuneda, and Teruo Okano, 「移植された培養ケラチノサイト細胞再生のための炎症性サイトカインの有効性」submitted to Journal of Investigative Dermatology
(6) Haruko Obokata, Masayuki Yamato, Satoshi Tsuneda, and Teruo Okano, 「ヒト線維芽細胞シートをルイスラットに皮下移植した後の超急性炎症の解析」submitted to Xenotransplantation.
(原文)
講演(海外)
(1) Haruko Obokata, Masayuki Yamato, Kohji Nishida, Satoshi Tsuneda, and Teruo Okano, "Subcutaneous transplantation of autologous oral mucosal epithelial cell sheets fabricated on temperature-responsive culture dishes", Society for Biomaterials 2007 Annual Meeting, Chicago, IL, April 17-21, 2007.
(2) Haruko Obokata, Masayuki Yamato, Kohji Nishida, Satoshi Tsuneda, and Teruo Okano, "Time-course observation of subcutaneous transplanted autologous oral mucosal epithelial cell sheets fabricated on temperature-responsive culture dishes",, The 13th World Congress of International Society for Artificial Organs, IFAO-JSAO Annual Meeting, Osaka, Japan, Jun 28-31, 2007
(3) Haruko Obokata, Masayuki Yamato, Kohji Nishida, Satoshi Tsuneda, and Teruo Okano, "Inflammatory アnalyses of subcutaneously-transplanted human epithelial cell sheets", Termis-AP, Tokyo, Japan, December 3-5, 2007
(4) Koji Kojima, Shohta Kodama, Haruko Obokata, Ana C Paz, Charles A. Vacanti. “Generation of Pneumospheres from Pulmonary Stem Cells.” Termis-NA, San Diego, December 7-10, 2008,
(5) Haruko Obokata, Masayuki Yamato, Kohji Nishida, Satoshi Tsuneda, and Teruo Okano, “Subcutaneous Transplantation of Oral Mucosal Epithelial Cell Sheets Fabricated on Temperature-Responsive Culture Dishes”, TERMIS-WC, Seoul, Korea, August 31- September 3, 2009
(6) Haruko Obokata, Masayuki Yamato, Kohji Nishida, Satoshi Tsuneda, and Teruo Okano, “Inflammation アnalyses after Subcutaneous Transplantation of Cell Sheets for a Novel Tissue Engineering Method”, TERMIS-WC, Seoul, Korea, August 31- September 3, 2009
(7) Koji Kojima, Haruko Obokata, Ana C Paz and Charles A. Vacanti. “Generation of Pneumospheres from Pulmonary Stem Cells.” TERMIS-WC, Seoul, Korea, August 31- September 3,2009
(8) Haruko Obokata, Koji Kojima, Masayuki Yamato, Teruhiko Wakayama, Teruo Okano, Satoshi Tsuneda and Charles A. Vacanti, “The potential of stem cells in adult tissues representative of the three germ layers.”TERMIS-NA, Orlando, December 5-9 2010
(9) Koji Kojima, Haruko Obokata, Jason Ross and Charles A. Vacanti. “Autologous Tissue Engineered Trachea with Epithelial Cell Sheets in Ovine Model.” TERMIS-NA, Orlando, December 5-9 2010
(和訳)
講演(海外)
(1) Haruko Obokata, Masayuki Yamato, Kohji Nishida, Satoshi Tsuneda, and Teruo Okano, 「温度応答性培養皿上で作製された自己口腔粘膜上皮細胞シートの皮下移植」, Society for Biomaterials 2007 Annual Meeting, Chicago, IL, April 17-21, 2007.
(2) Haruko Obokata, Masayuki Yamato, Kohji Nishida, Satoshi Tsuneda, and Teruo Okano, 「皮下移植後の口腔粘膜上皮細胞シートにおける再構築された超構造変化の時間経過分析」,, The 13th World Congress of International Society for Artificial Organs, IFAO-JSAO Annual Meeting, Osaka, Japan, Jun 28-31, 2007
(3) Haruko Obokata, Masayuki Yamato, Kohji Nishida, Satoshi Tsuneda, and Teruo Okano, 「皮下移植されたヒト上皮細胞シートの炎症分析」, Termis-AP, Tokyo, Japan, December 3-5, 2007
(4) Koji Kojima, Shohta Kodama, Haruko Obokata, Ana C Paz, Charles A. Vacanti. 「肺幹細胞からの肺炎球の生成」Termis-NA, San Diego, December 7-10, 2008,
(5) Haruko Obokata, Masayuki Yamato, Kohji Nishida, Satoshi Tsuneda, and Teruo Okano,「温度応答性培養皿上で作製された口腔粘膜上皮細胞シートの皮下移植」, TERMIS-WC, Seoul, Korea, August 31- September 3, 2009
(6) Haruko Obokata, Masayuki Yamato, Kohji Nishida, Satoshi Tsuneda, and Teruo Okano, 「新しい組織工学的方法のための細胞シートの皮下移植後の炎症分析」, TERMIS-WC, Seoul, Korea, August 31- September 3, 2009
(7) Koji Kojima, Haruko Obokata, Ana C Paz and Charles A. Vacanti. 「肺幹細胞からの肺炎球の生成」TERMIS-WC, Seoul, Korea, August 31- September 3,2009
(8) Haruko Obokata, Koji Kojima, Masayuki Yamato, Teruhiko Wakayama, Teruo Okano, Satoshi Tsuneda and Charles A. Vacanti, 「三胚葉各成体組織内在幹細胞の潜在能力」TERMIS-NA, Orlando, December 5-9 2010
(9) Koji Kojima, Haruko Obokata, Jason Ross and Charles A. Vacanti. 「羊皮症モデルにおける上皮細胞シートを伴う自己組織化工気管」TERMIS-NA, Orlando, December 5-9 2010
講演(国内)
(10) 小保方晴子、大和雅之、西田幸二、常田聡、岡野光夫、「温度応答性培養皿で作製した口腔粘膜上皮細胞シートの皮下移植」、第6回日本再生医療学会総会、横浜、2007年3月14日
(11) 小保方晴子、大和雅之、西田幸二、常田聡、岡野光夫、「温度応答性培養皿で作製した口腔粘膜上皮細胞シートの皮下移植」、第7回日本再生医療学会総会、名古屋、2008年3月14日
(12) 小保方晴子、大和雅之、西田幸二、常田聡、岡野光夫、「細胞シート皮下移植後の急性期炎症反応解析」、第29回炎症・再生学会、東京、2008年7月8日-10日
(13) 小保方晴子、大和雅之、西田幸二、常田聡、岡野光夫、「ヒト上皮細胞シート皮下移植後の経時的炎症反応解析」、第9回日本再生医療学会総会、広島、2010年3月17日
(14) 小保方晴子、小島宏司、大和雅之、若山照彦、常田聡、岡野光夫、Charles A. Vacanti,「三胚葉由来組織に共通した成体幹細胞の探索」第10回日本再生医療学会総会、東京、2011年3月1日-2日
国際特許
Haruko Obokata, Charles A. Vacanti.
Sub Population of Retained Embryonic Like Cells
(成体の中に存在する幼弱な表現系を有し、3胚葉系の細胞に分化することのできる幹細胞集団)
著書・著作物
大和雅之、小保方晴子 「成長因子」バイオマテリアルの基礎、日本バイオマテリアル学会、印刷中
- 2019/08/24(土) 22:49:55|
- 小保方さんの論文
-
-
| コメント:0































