AC129を巡る問題10からのつづきです。何かの文字と相性が悪いと書き込み禁止になるようです。L氏のコメントに対するこちらの応答に関する写真をここに貼っておきます。ここなら多分大丈夫ではないか。
自家蛍光というのはGFP以外の蛍光です。GFPの漏れ出しはGFPが光っている。この誤認に関して何か言ってる人もないですね。
>>
筆頭著者が不正をしないで誠実に真正なデータを残した場合でも、自家蛍光の誤認がそのままだと、Oct4-GFP偽陽性に基づいてキメラの実験に進んだと思われます。漏れ出しがあるのなら自家蛍光の誤認だと気づけないでしょう。デイリーも知らない事実でしょ。知らなければ漏れ出しは調べ方が適合的でなければ自家蛍光の中に含まれてしまう。だから考えが浅いと批判される。下の写真のOct4は内在性のOct4遺伝子が働いてOct4蛋白を作っているという証明で、Oct4-GFPが光ったからOct4遺伝子が働いたはずだという証明ではない。直接証明です。右端のGFPはローザクレですから細胞の由来を証明している。Oct4-GFPではないからOct4遺伝子発現の間接証明ではないが、実物が免染確認されているからそんな必要もない。以前L氏は両者の蛍光がずれていて同じ細胞ではないなどと言ってましたが、細胞塊はスライスされているからわずかにずれるんです。
この結果は丹羽さんのFigure3-bの結果と矛盾している。こんなにあっては変でしょ。同じ肝臓由来酸浴細胞塊です。要するに小保方細胞が何かということは完全にわかっているわけではないということです。こんなことはティシュー論文を読んだら最初からわかってることです。だからこそ研究しているんでしょ。小保方さんは正しく自分の細胞を再現している。キメラを捏造したのは若山さんです。最初は捏造のつもりでは無論なかったが、後の経緯でそうなってしまったということです。
(Ooboe さんからの連絡)旅行に出ている間に連絡がありましたので、ここで検討します。連絡事項は[Ooboeさん情報]に追加しています。ここにも再掲します。
>>
パートナーは、小保方stap研究を無きものにした画策を第一次、第二次、第三次の画策として分類した時系列説明資料を、検察に提出しています。
この中での犯罪事案は諸々存在していましたが、偽計業務妨害、背任、公務員法違反、公文書虚偽作成、告発幇助間接正罪などのうち、明確なもの、2件にしぼって、告発手続きでなく、申告書提出の手続きをしているところです。
検察からの文書も全てではありませんが、一部公開する予定です。担当官は公開okとのことです。
2019/11/27(水) 13:30:12 Ooboe
第一次共謀画策は、2014年3月~6月16日
第二次共謀画策は、6月25日~12月26日
第三次共謀画策を、2015年1月~6月
と大きく分類しています。
この経緯の中で、分水嶺時点を6月5日として検察に資料を集中して説明しています。
そのパートナー証拠の更なる裏付けをお願いしていますが、頑張れFBでは、詳細は控えるようです。
全経緯を通じての、解明ポイントを6月5日の理研本部会議時点として捉えると、6月25日以後の第二次共謀画策や第三次共謀画策の経緯の本質が浮上してくるので、犯罪事案の、間接的ではあるが、背景として、その内容を検察に理解して貰うのに焦点をあてたとのことです。
この詳細は、ブログ開設で展開したいそうですが、検察には、この首謀者と共謀者を証拠をもって特定して提示しているとのことです。
この首謀者については、居士さんの考察とは異なるものとなっています。
第一次画策以前の2011年から2015年の第三次画策の全経緯を通じて一貫した首謀者としてパートナーは検察に提示して説明しています。
抽象的で、ごめんなさい。
2019/11/28(木) 00:33:12 Ooboe ①小保方stap研究を無きものにした画策
②共謀画策は三次にわたる
第一次共謀画策は、2014年3月~6月16日
第二次共謀画策は、6月25日~12月26日
第三次共謀画策を、2015年1月~6月
③分水嶺時点は6月5日の理研本部会議時点
④犯罪事案は以下の内の二件に絞って申告書提出手続き中
偽計業務妨害
背任
公務員法違反
公文書虚偽作成
告発幇助間接正罪、等
纏めると以上でしょうか。6月5日の理研本部会議時点がエポックであるということは会議に参加していた人たちの中に首謀者、もしくは首謀者にそそのかされている人が潜んでいるということになりますね。我々の今までの交換情報の中では松崎氏ということになると思います。
「この首謀者については、居士さんの考察とは異なるものとなっています。」の意味が二通り考えられるので、このことに関して少し述べておきましょう。
私は"犯人"は若山さんと言ってます。この意味は明確で、キメラができたのは若山さんのntES化手技の結果で、論文に書かれた小保方細胞の自然の結果でもなければ、小保方さんのESコンタミ捏造でもないという意味です。その意味で、私は犯人は若山さんだと言ってる。
対して、まず最初に、パートナー氏の(Ooboeさん、和モガさん、Ts.Markerさん、学さん、アルイミオウジさんをも含むかもしれませんが)推測の前提として明確に私のストーリーと対立しているのは首謀者ではなくて、論文通りにキメラのできているSTAP細胞があるか無いかという点です。
①論文通りのSTAP細胞由来キメラがある。
②小保方さんがESによって若山さんに捏造させたキメラがある。
③若山さんが小保方細胞核をntES化したものからのキメラがある。
パートナー氏の推理は①です。私は③です。桂報告書は②です。
②はありません。パートナー氏も私も否定済みですね。ため息ブログや雑談コーナーももはや小保方さんの論文不正を言い立てるばかりで、なぜキメラができたかに関しては口を閉ざし始めていますね。あそこにたむろしているのは小保方捏造を騒ぎ立てたマスコミ関係のスピン屋たちばかりです。言い訳集団です。医学界関係の論文不正というのは昔から有名でね。医者が自分の箔付けのために博士号を欲しがるので、一回限りの論文ということで審査や内容はユルユルで、不正は昔からまかり通っている。細胞生物学会にもはびこっているんですね。だいたい学会というのは利権団体ですからね。言うまでも無いが立派な医者、立派な研究者も昔から少数ながら常に存在している。そうでないと社会は維持できない。
小保方さんの不正の中で一番重大なのはメチル化検証です。これは若山さんがキメラを作る前からもちゃもちゃいじっていてプログレスレポートでいい加減な報告をしている。これは若山さんに誤解を与えたかもしれません。ただし、三誌論文にこのデータを使ったというのはキメラが出来ているんですから、若山さんが誘因になっている不正です。無論、不正は不正で、そもそも研究というのは真実を知るために実験しているのであって、実験事実に忠実でないものは、研究姿勢に既に問題がある。モラルなんて問題じゃない。ましてお作法なんて落第生に言う低レヴェルの話だ。意図しない結果を実験が教えてくれているのにそれを無視するなんて研究者じゃない。心が鍛えられていない。もしくはふんどしが緩んでいる。
①は再現実験ではできなかったので論文発表が早すぎるということは明確にされたが、あの再現実験は制約がありすぎますし、何をしたかに関して、隠されていることが多すぎる。特に三胚葉分化実験をしたかしなかったかに関してはとても深い疑義がある。その意味でできないということが証明されたとは言えません。でも、あり得た場合になぜ、理研が世界に向かって記者会見発表までしたものを後に無かったことにしたのかの説明が必要です。誰が、できているものをできていないことにして隠そうとするのかを考えないといけない。
この結果は世界的に大きな利権になるから理研が隠そうとするということはあり得ませんね。世界に向かって記者会見したのは理研です。
もうしてしまってから、初めてそれを知って、理研に対して隠せと命令できるとしたらもっと上の組織で、官僚組織ということになるが、これもないですね。笹井さんは文科省との折衝役で事前に文科省に説明しています。小保方さんをつれて行ったこともあると手記には書かれている。記者会見発表前には文科省からの天下り役員も知っていて連絡していますね。こういうのはないです。
「小保方stap研究を無きものにした画策」という言葉の中にSTAP研究と書かれているが、この研究はキメラができたことを事実として含んでいるものなのか、それとも理研以前の段階、つまりティシュー論文の研究までを意味しているのが曖昧です。
後者であっても現実的には小保方さんの博士号を剥奪し、ヴァカンティと小島両氏の地位を失わせたのですから、"無きものとした画策"ということはできます。どちらなのか、どちらもなのか、明確にしたいものです。
ただ、この課題は無数にある科学的課題の中の一つに過ぎません。時がたてばこの研究に興味をもってやり直す人は出てくるでしょうから、隠さなければならないほどの利権を生み出す課題でもないですね。利権になるのは臓器にまで作れた場合で、ESですらとんでもなく遠いのですから、こういう基礎研究を隠すなんてこともありません。笹井さんを失ってもっと遠くなってしまったとも言えますね。でも、弟子が続くでしょう。
[①論文通りのSTAP細胞由来キメラがある。]のに、なぜ隠そうと画策するのか。大した利権でないのに、隠そうとした動機は、だから[論文通りのSTAP細胞由来キメラがない]からだと、私は考えたんです。
私の方に来るためにあなた方をそちらに引き留めている理由をちゃんと書かないといけない。逆にその理由に私が納得したら、私の方からあなた方の方に行くでしょう。
私はあなた方が意図する意味での理研内の首謀者としては松崎氏であると考えている。笹井さんに対する内部権力争いにこの問題を利用して、報告書の基礎になる実験結果を提出した。ただし、ここには早く収拾しろという文科省の圧力もあって、利害が一致したということはありそうですね。もう一人松崎氏以外にGDがいますね。そちらですかね。そこは私は分からない。
無論、私は松崎氏を誘導したのは若山さんだと思っています。嘘をついて騙している。松崎氏はそれを信じて実験を行っている。ESコンタミ捏造だという結論を想定して実験を行っているんです。報告書が若山さんを全く容疑者として扱ってないことは自明ですね。全く客観的な調査ではない。
私のストーリーが正しいとして、後の若山さんの行動から人々は若山さんを疑えたかと考えるべきです。結構上手に騙しているんです。そもそも科学者はこんなことで人を疑いませんからね。私のストーリーに思い至れない人々は小保方さんのESコンタミとしか思えませんよ。たくさんそうでない事実を見ているんですけどね。若山さんがESコンタミ捏造なんてするわけはありませんからね。すると意図しない経緯でこうなったという事情は気づいていませんから調査できません。こういうことにはプロの捜査に慣れている警察を入れないといけないんです。業界の自主運営ではいけません。欧米は皆そうなってる。
(学さんの誤解)序なので、2019/11/28付の学さんの考察記事に関してコメントしておきましょう。誤解がありますね。以下です。
>>
年数が経つと、1塩基変異が積み重なってきて、コロニーごとのマウスのSNP(1塩基単位の突然変異を含む)が、離れてくる現象について議論してます。129であれ、B6であれ、その近交系マウスに特異的SNPsは100万か所単位で存在しています。調べているのはその中の一部だけです。まして、登録されていない新たにできたSNPsなんて調べられてはいません。
Ts.Marker さんのブログの2015/12/31の記事に以下のようにある。
>>
------ 追記 6/30 -----
マウスのSNPsは1000万といわれていますが、調査報告ではB6と129で違う300万を使用している。(あのねさんへの返事)学さんのブログにあのねさんがコメントされているのに気づきました。以下です。
>>
あのね
このコメントを参考に考えて見ましょう。一言さんもブログで、なぜRosa?と書かれたようにブログで追加ご考察して下さい。
>5603. L
2019年11月17日 06:58
モンキーさんのコメントに納得できないのは、あまりに笹井先生に失礼な推測だと思うからです。笹井先生がそんないい加減な形でメソッドを書くとは考え難いです。なので、筆頭著者が書いた(あるいは筆頭著者からのインプットで笹井先生が書いた)と私は推測します。この場合は、まず、筆頭著者に誰かからRosaマウスについてのインプットがある必要があると思いませんか? だとすると、どのような意図でインプットがあったのか、疑問に思いませんか? 謎でしょ?
Rosa26の遺伝子座にGFPを挿入したマウスは、理研CDB相澤先生が動物資源開発センターの室長兼任で当時、確立してして論文発表しています。アブストを貼り付けますので日付に注目して下さい。2011年7月前後の時期に若山研究室ではどのようなSTAP実験をしていたか想像しましょう。
丁度、若山研でGOF-ESを作って、後に2つの近交系129マウスとB6マウスを交配させてF1マウスを作ろうとしていた時期だと思います。この時期はすでにCDB内部で129 carrying Rosa-26GFPマウス完成の案内通達があったはずで、研究所内で都合の良い129GFPマウスが配布されているのに、市販の129マウスだけを買う人はいません。若山氏はこのマウスも使ったと思います。後にこのマウスを調べると困る流れになります。FES1の由来のマウスに合わなくなるからです。そしてそんなマウスは無いことで済ませて、遺伝子配列は別口ですでに登録されているのにゲノムの比較調査されていません。
http://www.cdb.riken.jp/jp/04_news/articles/11/111212_liveimagingmice.html
独立行政法人 理化学研究所 神戸研究所 発生・再生科学総合研究センター
2011年12月12日 「ライブイメージングに適した新たな蛍光マウスを開発」
発生過程において組織や器官が形成されていく仕組みを知るためには、個々の細胞の振る舞いを調べる必要がある。蛍光ライブイメージングは、生きたままの胚において特定の細胞や細胞内の構造を可視化し、その動きを追跡することができる技術である。マウスにおいても蛍光ライブイメージングが可能だが、多くの場合、蛍光遺伝子を染色体にランダムに導入しているため、「←ここ重要」発現部位を厳密に制御できない、発現量にバラツキがある、複数の構造を同時に標識できないなどの問題を抱えていた。理研CDBの動物資源開発室(相澤慎一室長)は、核や細胞膜など7種類の細胞内小器官を条件特異的に蛍光標識できる12系統のマウスを開発した。ライブイメージングに適した十分な蛍光シグナルが得られ、また、二重標識も可能であることが確認された。既に汎用されているCre-loxPシステムを発現制御系に用いているため、容易に発現部位を限定できる。この研究成果はGENESIS 誌の7月号に掲載され、同室はこれらのマウスの配布を開始している。
これが129/Sv carrying Rosa26-GFPマウス。ところが、桂報告書10P
「…なお、Article のメソッドに、129/Sv carrying Rosa26-gfp からキメラ寄与能を有する STAP 幹細胞が樹立された、との記述があるが、129/Sv carrying Rosa26-gfp マウスは理研 CDB に導入された記録や飼育記録はないことから、これは誤記と考えられ、若山氏の説明によればここで言及された STAP 幹細胞は AC129である可能性が高い。」
CDBに導入された記録や飼育記録はないなんて、よくも平然と言えたものですね。「これは誤記と考えられ」と含みを残して「誤記である」となぜ断言できないのですかね。少なくとも小保方さんが笹井先生らにSTAP細胞のライブイメージングで見せて「良く光っているね」と感想をもらしたマウスはRosa26-GFPマウス(若山研B6と交配F1を含む)のSTAP細胞であるほうが自然かなと思います。自己保身で調査に率先して協力した=小保方研に都合の良い細胞だけを残し、泥棒細胞を抱えたオフサイド若山さんの言質を取った調査委員会は市販の129マウスだと信じた。確証バイアスは恐ろしい。CDBで配布されたマウスも記録なしとなる。STAP論文が取り下げられて、なおかつ追撃でSTAP現象すら否定なら「嘘も方便」とはこのことですね。
調査報告書でも指摘されていたように、若山さんのマウスコロニーの管理の無頓着さについては、「あの日」の記述から考えられる考察案件を持っており、追ってそもそも論から後にコメントします。
2019/11/29 URL 編集この件は既に考察済みなんですが、あれこれと何度も考え続けていますから、人にはなかなか分かりません。再説しますと、この相沢さんのマウスは6種の色違いの蛍光遺伝子とクレロックスシステムを使って最終的に12種類の同時識別可能なレポーター遺伝子を造れたということです。その時に使われたマウスの背景はここには書かれていません。従って、129/Sv carrying Rosa26-gfp マウスは無いと言われたらそのまま受け取るか、無いという証拠を出せというよりない。誰もその試料を開示請求していません。
私が今考えているのは誰かが小保方さんに129/Sv carrying Rosa26-gfp だと言ったから、又誰かがキメラを作ったと言った、もしくは言ったと誤解したから、彼女は論文にそう書いているということです。
そもそもなぜ若山さんがこんな時期に背景マウス種の違いによるSTAP細胞の樹立確認をしたのか、しかも後にキメラは作らなかったと言っていて、確認目的ならキメラまで作ってみないと変なので、これは目的が違っていて、TCR再構成結果に対する別の関心から小保方さんに129のSTAP細胞を作らせたのではないかという可能性を今、考えているんです。
(和モガ氏のヘテロプラズミー)Ts.Marker さんがいろいろと調べていましたが、最終的にNOD / ShiLtJまで突き止めてその後の進展が無かった。このマウスは免疫不全マウスなので、このようなマウスはクローン作製には使われない筈なので変だなと思考停止していました。この件に関して和モガさんも考察を続けられていたのはずっと読んでいましたが、今又読み直して読み落としに気づきました。以下のように書いている。
>>
NOD / ShiLtJマウスを提供しているCharles River社のホームページにはNOD / ShiLtJマウスは非近交系のICRマウスが起源であると書かれている。ICRは若山研でクローン胚を作るときのマウスです。ただ、Ts.Marker さんはICRを調べてない。
>>
-------------- 12/7 調べ -----------------------
Mouse strain assembly hub - May 3, 2017 より
DBA/2J T (https://www.ncbi.nlm.nih.gov/nuccore/CM004183.1)
129S1/SvImJ T (https://www.ncbi.nlm.nih.gov/nuccore/CM004179.1)
BALB/cJ T (https://www.ncbi.nlm.nih.gov/nuccore/CM004214.1)
C57BL/6NJ T (https://www.ncbi.nlm.nih.gov/nuccore/CM004277.1)
NOD/ShiLtJ A (https://www.ncbi.nlm.nih.gov/nuccore/CM004185.1)和モガさんの整理しているものもここに貼り付けて置きましょう。分かりやすいです。
ICRのデータがAであれば、私の小保方細胞核使用ntES論にとっても強力な傍証になるでしょうね。ただし、ヘテロプラズミーは必ず起こるというものでもないんですね。
(学さんの誤解2)まだ学さんの誤解が続いているようです。ため息氏とのやり取りですが、ため息氏の認識が正しく、学さんは勘違いしている。これで双方で悪口の言い合いをするので、小保方さん擁護の学さんの旗色が悪くなるから、世間はスピン屋たちが正しいと誤解する。私はこれをスピン屋同士の芝居だと見做したわけですけど、Ooboe さんはそうではないとおっしゃるので、学さんの誤解だとして指摘し続けなければならないようです。
>>
F1では、129とB6がランダムに混じりますので、子どもごとに違う遺伝子構造ですが、129B6ES1~6の例でもわかるように、両親がそれぞれ、同じ遺伝子構造なので、その子どもごとに129とB6が混じり方が違っても、それでも、子どもの遺伝子の類似性が高いことがわかります。説明の仕方が悪いんですね。「F1では、129とB6がランダムに混じります」というのは近交系が崩れている時の話ですよね。先に、完全な近交系ではランダムには混じらないという原理から順番に書かないから支離滅裂になるんですね。完全な近交系が保たれていたら「子どもごとに違う遺伝子構造」にはならない。何匹生まれても全部同じ遺伝子構造のF1になる。
それを説明した後に、マウスコンタミがあって近交系が崩れている場合は「F1では、129とB6がランダムに混じりますので、子どもごとに違う遺伝子構造」になると説明したらいいんです。
最初に説明すべきことを誰でも知っていることという前提で端折るだけなら、少しずつ違うのは当然だということで分かる人には誤解が無いんですが、逆になぜ少ししか違わないのかという説明として、後から、誰でも知っていることとして端折ったはずの説明を加えるから、あなた、どういう理解をしているのと、分かる人から反論されることになる。
>>
F1の場合は、それぞれ近交系で、129コロニー、B6コロニーでは、ほぼ同一遺伝子状態であっても、交配してしまうと、できてくるF1マウスは、それぞれ129とB6の混じり方が異なってくる。ここも、「交配してしまうと、できてくるF1マウスは、それぞれ129とB6の混じり方が異なってくる。」のは近交系が崩れている場合ですから、前の文章に「F1の場合は、それぞれ近交系で、129コロニー、B6コロニーでは、ほぼ同一遺伝子状態であっても」という説明を置くと前後の文章が矛盾してしまう。「ほぼ同一遺伝子状態」が「同一」だと言ってるのか「同一でない」といってるのかというと、「も」が入っていることによって「同一」だと言ってることになるんです。作文の仕方が間違ってる。日本語の「も」の使い方の間違いです。学さんは日本人ではないのかな。
>>
2019年12月2日 4:12 PM
>学とみ子は両親がヘテロザイゴート のとき作成される配偶子は均一にならないからその子供同士のSNPは一致しないのがわからない?
近交系というのは兄弟同士等を掛け合わせて、すべての遺伝子(DNA)をホモザイゴートにしたマウス のことでしょ?ホモザイゴートだと減数分裂にのとき、交叉が生じても配偶子の遺伝子構成は変わらないから兄弟のSNPsはほぼ一致する。ため息さんの頭の中には、過去に129とB6が混じってしまった話と、自然に生じる塩基変化で遺伝子型が変化していく話とが区別つかず、混乱しているようです。
ホモザイゴートとは、129のみで維持、B6のみで維持の場合ですよ。事故的に他のマウスが混じっても、又、ホモザイゴートのコロニー内で、マウスの遺伝子は変化していきますよ。①完全に近交系マウス同士のF1は何匹生まれても両親から半々に遺伝子を受け取るので完全に同じ遺伝子構成になる。子供同士でわずかずつ違うということはない。
ため息氏は近交系の基本認識を言ってるだけです。野生のマウスは両親の遺伝子を受け継いでいるので減数分裂の時にできる配偶子は遺伝子の並びが引き継いだ両親の遺伝子に分かれるので2種できる。でも、近交系のマウスはその両親が同じだから減数分裂しても全く同じ配偶子ができる。だから129とB6を掛け合わせた子は何匹生まれても遺伝子構成が全く同じ混血の子供たちが出来るんです。混血ではあるが子供同士で遺伝子の並びに違いがあるということはないんです。この時点でため息氏は何も間違ってない。
②マウスコンタミがあって近交系が崩れていたらどうなるか。
既に論じたように、若山研でのマウスコンタミには二つの要素があって、129に関してはそもそもジャクソン研究所で古くにB6の飛び込みがあって気づかれていなかった。
もう一つは、若山研で自家繁殖させている間に生じたマウスコンタミがある。
SNPsに関しては前回指摘したように、調査されているSNPsはそれぞれの近交系マウスの特異的SNPsとして登録公開されている1千万個程度の中の、特に、今回では129とB6で違っているSNPsの3百万個だとTs.Marker さんが書いていると指摘した。
SNPsはマウスを飼育していると生殖細胞にも突然変異が起きてきますから、最初に登録されていた1千万個に対して継代ごとに何個か、何十個かずつ増えてくる。でもこれは最初の登録されている場所とは違います。1千万個に対して有意なだけ増えると別種の近交系マウスとして新たに特異的SNPsが登録されます。それまでは変更ありません。学さんは新たに増えたものを調べていると思われているが、そんなところは調べられてない。
次にSTRに関してはFES1とFES2で違っていたからFLS3はFES1と同じだと東大グループと遠藤氏が結論したと日経サイエンスに書かれている。我々は無論若山さんがFES1のチューブにFLS3を入れたからだと思ってますが、それはともかくとして、同時期に同じ近交系マウス同士の親を交配して作ったらしきFES1とFES2のSTRが違ってるなんてありえないと知らねばなりませんね。
FES1とFES2のSTRが違いうるという説明を太田さんにして欲しいね。どんな交配したの。③培養変異の存在。
これは体細胞分裂ですからね。別の話です。
さて、学さんの「も」の使い方は変なんです。「ホモザイゴートとは、129のみで維持、B6のみで維持の場合ですよ。」と書かれているのは近交系なら当たり前のことです。そしてこのことをため息氏が言ってる。言ってることが同じならここで又確認する必要は無いでしょうよ。そこにどうして、「事故的に他のマウスが混じっても、又、ホモザイゴートのコロニー内で、マウスの遺伝子は変化していきますよ。」という訳の分からない文章が続くのか。
「事故的に他のマウスが混じっ」たら、既に近交系マウスではなくなっている。それはここには引用してないがため息氏は分かっている。それは向こうのブログに書いてありますね。ここで学さんが引用しているため息氏の文章の近交系マウスの認識は正しいんでしょう。そういう場合はまず正しいと認めた上で、私が言ってるのはマウスコンタミのことだと言えばいいだけでしょ。私の言いたいことはそこでは無いと。
でも、「事故的に他のマウスが混じっても、」という文章はどこに落ちていくんでしょうかね。事故的に混じったら近交系ではなくなる。はい。で、「又、ホモザイゴートのコロニー内で、マウスの遺伝子は変化していきますよ。」とは何を受けている文章なのでしょうか。これって「事故的に他のマウスが混じったら、又、ホモザイゴートのコロニー内で、マウスの遺伝子は変化していきますよ。」の間違いじゃないの。「も」が入ると意味が変わる。日本人じゃないんですかね。外国人ならこういう間違いは理解できますけどね。
学(ガク)さんって本名なら珍しいお名前ですが、ひょっとしてシナ系の帰化人の方ですかね。もしそういう帰化人1世のかたであったら、このことはとても理解可能ですね。そしてそういう配慮で読むことができる。
「も」の使い方の指摘は2度目です。なんとなくそうではないかと思えてくる。
ため息氏のスピンはそんなところにはないので、こういうやり取りをされて、本質的でないところで学さんがやり込められると、小保方さん擁護派は困るんですがね。
(アルイミオウジ氏の最近の動向)アルイミオウジ氏を常にチェックしているのは最近では私くらいのものではなかろうか。
上の学さんの相手が自分のブログのコメント欄に書いているのが以下です。
>>
sigh
2019年11月22日 9:21 AM
>学とみ子
擁護の阿塁未央児さんも「学さんは何が言いたいのだろうか? FES1とFES2で雄親のコロニーが違うという示唆が、桂報告書のどこにあるのだろうか?」と呟いていますよ。
何が言いたいの分かっている方はいないようです。説明したらどうでしょ?ここで紹介されているアルイミオウジ氏のツイートが以下です。
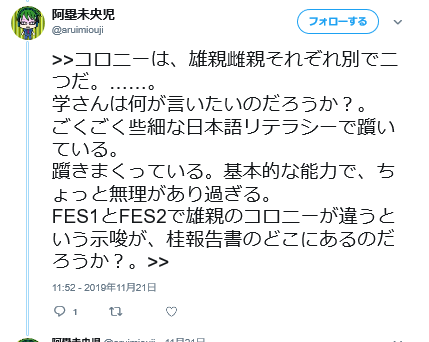
学さんが言ってるのは上述したように同じコロニーから作られた筈のFES1とFES2のSTRが違ってるのはなぜかということでしょうね。
このこととは別に最近アルイミオウジ氏が面白いことを発言している。以下です。
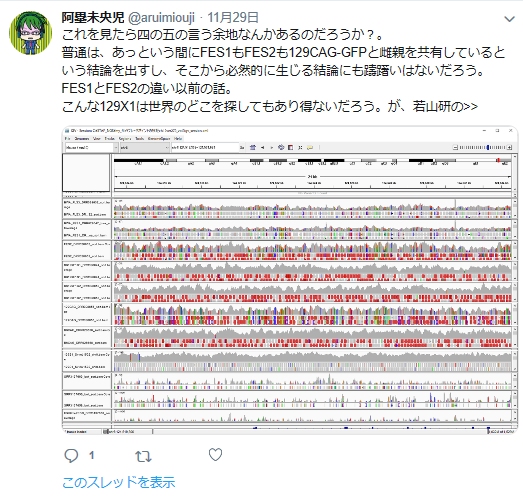
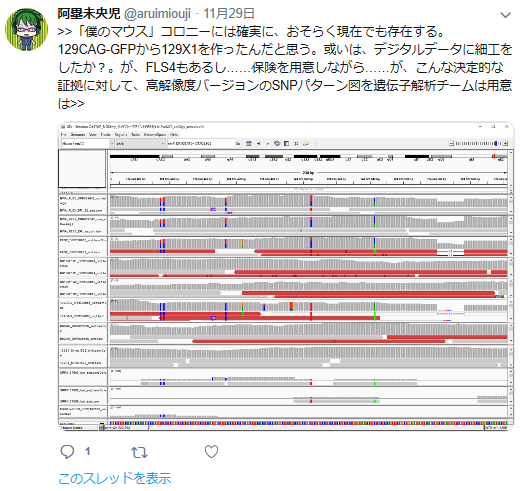
私の疑義していたことですね。
>>
AC129を巡る問題8
(残された問題の整理)
さて、問題の数も少なくなってきましたね。ただし、解の困難さは増してきました。ここらで残されている問題を整理しておきましょう。
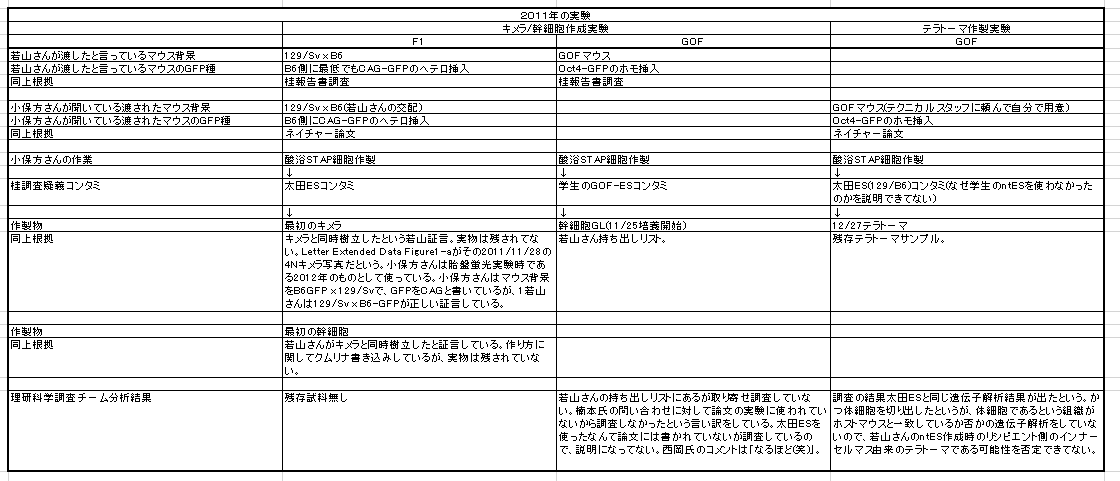
①FLSは「僕のマウス」を渡したと一方的に証拠の提出もなく若山さんが主張しているが、実際の検査で出て来たのはAcr-CAGのB6(岡部マウス)と市販のGFP無しの129X1/SvのF1だという報告書の調査結果となっている。しかし、129側の18番染色体に1コピーのCAG-GFPが入っているのではないかという疑義がでた。なぜなら、SNPs解析結果がこの129は「僕のマウス」の片親であることを示唆しているからだ。最初に放医研がFLSを検査した時、15番にヘテロに4コピーのCAGがあり、18番にはGFPが無かったと発表していたが、後に15番のGFPが否定されたが18番に関しては何も触れられなかった。又間違いの原因もちゃんと説明されていない。内在性アクロシン遺伝子を間違えたのならホモだと思うはずでヘテロに間違うというのは考えにくい。調査自体がとてもあやしい。すると18番に無かったというのは本当なのかという疑義が残るということです。理研のチームはGFPを自分でちゃんと確認したかということです。この説明も無い。アルイミオウジ氏は学さん以上に説明能力の欠如している人なのでどういうことをしているのかということがちゃんと確認できるまでは何とも言えませんが、129CAG-GFPマウスから129X1を作ったという説明は聞き捨てならないところです。これは129X1に戻し交配したということですが、129X1があるのに戻し交配してGFPを消すという意味が分かりません。これはGFPの有無を調べればいいだけのはずですが、GFPは無かったのでしょうか。ちょっと明確でないところです。ただ、129にB6のコンタミしているSNPs分布が「僕のマウス」の片親である129と同じだという分析結果は私と同じですね。
また最近彼がやってるのはSNPsでなくてSNVsを見ているらしい。頻度が1%以上ならSNPであるが、それ以下まで入れると無数にあることになる。何をやってるのかちっともわからない。以下も、勘違いということでいいんだよな。
 (学さんの誤解3)
(学さんの誤解3)学さんが何を誤解されているのかが段々はっきりしてきましたね。
>>
近交系マウス由来同士に、予期せぬ交雑がおきると、別の系統のSNPが、マウスに入ります。
さらに、交雑が進むと、別の系統のSNPをホモでもってしまうことがあります。
かつて混じったというと証拠でしょうが、近交系コロニーに戻して年数が経つと、次第に別の系統由来SNP変化が少なくなり、ワトソンさんらは、薄まると表現されたと思います。
以前に、専門家の方が、ヘテロはホモに収束するとか書ていたのを見ましたが、細胞は独自に、SNPの修復を行うと考えて良いのかな?、ここは確かでないです。近交系マウスは基本自家繁殖させてはいけないです。マウス供給業者との契約でそうなってるはずです。
近交系マウスは兄妹交配を続けます。親にはかけ戻しません。親の管理ケージと生まれた子供たちの管理ケージは分けられています。
兄妹交配した子供は親たちよりホモ率が高いんです、その原理を使って20継代以上させて98%以上のホモ率になったものを近交系マウスと称して業者が販売している。
その後、買ったマウスの雌雄を交配させて維持繁殖させるときに、業者と同じように兄妹交配管理を続けているとホモ率はどんどん上がっていきます。以前そのデータを貼り付けましたね。
でも、買ったマウスをどのような管理をするのかは研究者たちの自由ですが、自分で繁殖させたものを使って論文を書くときに購入業者の名を使うと違法になりますね。論文にマウスの業者を特定させたいときは買ったものそのものを使わないと業者は責任取れません。今回みたいにこれだけのマウスコンタミがあると研究目的によっては責任問題になりかねない。間違った取説で使用されたものにメーカーは責任取れません。
若山さんのところではいろんなマウスを自家繁殖させていました。クローンの研究者ですので、作られたキメラの貢献度を見るためにGFPが入っていればいいんですね。基本自分たちの研究目的に支障が無ければある程度のコンタミは生じていても構わない。
そういう環境の中で今回の事件が発生している。小保方さんの研究は若山研のマウスで行われている。それ以前は彼女は自家繁殖マウスは使用していません。業者から買ったものだけです。
もし実験が購入マウスだけで行われていたら細胞の由来トレースはとても困難なものになっていたでしょうね。
若山さんがどういうマウス管理をしていたのかは分かりません。自家繁殖させていたらしいのは以下です。
①GOFマウス
②ロックフェラー大学で作ったB6-CAG-GFPホモマウス
③②からGFPを移し替えた129/Sv-X1-CAG-GFPホモマウス
④太田さんが岡部研から持ち込んだB6-Acr-CAG-GFPホモマウス
他のものは情報が無いのでわかりません。動物実験申請書では業者名が黒塗りされていて、恐らく自家繁殖分と書き分けられているので黒塗りされているのではないかと思われますが、詳細が分からない。
①は小保方さんが手記でばらしてしまった。②③は「僕のマウス」の嘘の言い訳をするために記者会見で自分でばらした。④も記者会見で岡部さんに譲ってもらって維持していたとばらした。手記では顕微授精もしていたような書きざまになっている。自家繁殖させているとお金の節約にはなります。
どの自家繁殖も業者のように兄妹交配を厳密にやっていればホモ率はどんどん上がってくる。でも自家繁殖でしかも研究目的からしてそれほど関係のないものなら、一つのコロニーで維持繁殖させる。すると近交系マウスから所謂クローズドコロニーマウスになる。親との戻し交配も発生しますから近交系マウスの原理通りにはホモ率は上がりません。それでも買った時の近交系は維持されています。
ここに違うマウスのコンタミが入るとどうなるかということです。白毛の129のコロニーの中に黒毛のB6が飛び込んで気づいて取り除いた時には既に交配していたとする。その子は全部混血色ですから生まれた時に取り除く。これでコンタミは取り除かれた。でも、いつ気づくかによりますね。生まれた子供が更に交配するほど長く気づいていないと、F2では白毛の子もいくつか残る。すると毛色以外の遺伝子がB6である白毛マウスが残ってしまう。マウスが成熟するまで1か月以上はかかりますから、こういうことはとても起きにくい。
今回のコンタミで顕著なのは③のマウスですね。B6から129へGFPだけを移し替えた。その手法は基本的にF1に対する129の戻し交配です。半分B6が入ったものでGFPの光っている子供だけに129を何度も交配させる。これも20継代続けると近交系になります。若山129近交系GFPマウスです。このマウスに調査時点でB6のSNPsが所謂近交系マウスとしては大量に入っていた。入っていたのは事実で、しかし、この原因が分からないわけです。
①戻し交配回数不足
②そもそものジャクソン研究所でのコンタミ残滓
③若山129近交系GFPマウス樹立後の新たなB6の飛び込み事故
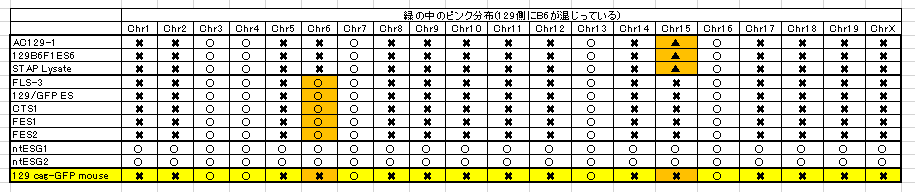
もう一度確認しましょうかね。BCA報告の図です。
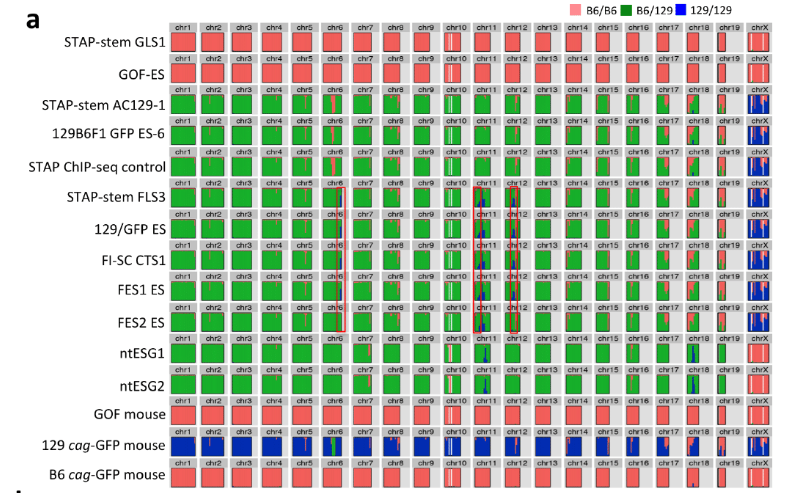
まず、ntESG1、G2の赤を見てください。ほとんどありませんが僅かに7,10,16番にあって他の細胞とも共有されている。これが②のジャクソン研究所でのコンタミ残滓ではないかと推測される。この129はterでテラトーマ専用に開発された129の亜種です。
それに対して、129cag-GFP mouseは129のX1です。市販のX1はデータとして示されていません。若山さんの「僕のマウス」の片割れの親マウスのデータだけが示されている。このコンタミはひどいですよね。市販されたものにこれだけコンタミがあるということはあり得ませんので、このコンタミは若山さんが作ってしまったものです。
一番特徴的な17,18,19番染色体を見ると上から順にAC129からFES2まで8ライン全部に共通していますね。
上3つは「僕のマウス」ESで捏造したと言ってるわけですから、一致しているのは分かりますよね。でも、更にFLSから下三つは太田ESで捏造したと言ってるんでしょ。更に下2つの太田ESはアクロシン入りの岡部マウスとのF1ですけど相手の129は市販のGFPのない129じゃなかったですか。というのも彼は最初自分はterと交配したつもりだったと証言していて、加えて、自分はB6に関してはアクロシン入りの岡部マウスしか使わないのでただ単にGFPと書いたと日経サイエンスに証言している。129側にもCAG-GFPの入った「僕のマウス」の片割れ129を使ったのならGFPに関してそれと注記するでしょう。
FES1、2のB6コンタミが他のラインの129側のコンタミと一致しているのは若山さんがFES1,2にFLS3を入れ替えたからです。私がなぜアルイミオウジ氏の最近の動向を報告したのか。このことがあるからです。彼の報告にGFPの有無検証が出るのを待ちましょう。
疑義は以下の解析は間違っていのだが、正しい答えは示されていないということです。

放医研は何を探していたのでしたかね。
CAG-GFPがどこに有るかですよね。放医研は最初15番にヘテロで入っていたと報告したことになっている。
でもそこにはGFPは無かった。実際には3番染色体にあったのです。岡部マウスのAcr-CAG-GFPは3番に有るんです。
ではどうして放医研は15番染色体にあると言ったのか。3番に有るのは内在性アクロシンプロモーター配列です。
では彼らは若山さんからプロモーター配列を知らされていてアクロシンプロモーター配列を含むプライマーで探していたのだ。若山さんは放医研が岡部マウスのGFPを見つけてくれると期待していたのに放医研は内在性アクロシンのプロモーターを捕まえてしまったのです。
ということはここにある18番にGFPが無いという結論も間違っている可能性がありますね。放医研はCAG-GFPを探しているのではなくてアクロシンGFPを探している結果になっている。18番にアクロシンプロモーターが無いのは当たり前です。ではCAG-GFPは調べ直したか。彼らは3番にAcr-CAGがあったと訂正した。では18番にヘテロで1コピーCAGが入っているか否かを調べたか。
少なくとも報告書は沈黙している。我々のSNPs解析の分析では129は「僕のマウス」の片割れであるという結果になっている。つまり18番にヘテロで1コピーのCAGがあるはずだと。
上の記者会見の説明映像のFLSはアクロシンプロモーターを探した結果です。右のAC129等はCAGプロモーターを探している。プライマーが違うんです。もしCAGプロモーターでFLSを調べていたら15番なんかに見つかるわけがない。そこにはCAGはありません。犯人は若山さんですよね。
(学さんの誤解4)今度は、
「すり替え、入れ替えなど、個人的な犯罪的行為をできるだけ考えないで、誰でも納得のいく説明はないものか?」という考察のようです。そこは最初に考えるところでないといけませんよね。まだだったんでしょうか。
互いに主張している事実関係に関して矛盾があるとどちらかが嘘をついていることになるんです。
ここをまず調べてないというところが怠慢ですね。
若山さんは「僕のマウス」を渡したと言っている。小保方さんは渡されたマウスでSTAP細胞を作って返したと言ってる。出来ている幹細胞を調べたらアクロシン入りだった。
若山さんが嘘をついているか、小保方さんが嘘をついているか、第三者が何かしたかのいずれかです。
この構図に関して一度も考えたことが無かったのでしょうか。
最初のキメラに第三者がESを入れて成功させてやるなんてことがあるでしょうか。いろんな種類の背景マウスですべて若山さんが渡したと主張しているマウス背景でない検査結果が出ている。第三者があり得ない決定的な証拠は最初の捏造が成功している捏造だからです。どうして成功していない段階から成功させるような手伝い捏造コンタミをする第三者がいますかね。この第三者は始めに成功させるようにコンタミさせた。ということはそれ以降も目的は成功させるようにコンタミさせていたのだということになる。どこにそんな第三者がいますか。
犯人は若山さんか小保方さんしかいないんです。そのことをまず押さえないと。選択肢が無くなったところから、ではどちらなのだと考える。
若山さんが嘘をついているのなら、彼の言ってることは一つも信じられない。小保方さんが嘘をついているのなら彼女の言ってることは皆嘘だ。二つを比較したらいいんですね。
彼女は手記でたくさんの証言をした。再現実験にも参加した。そして言ってることには嘘が無い。対して、若山さんは実験にも参加せず、矛盾に対する説明が無い。そして既存ESコンタミだとしている桂報告書がすべてだと証言した。つまり、既存ESがどこから来たのかということを証明しないと彼の証言は一つも信じられない。パートナー氏の質問状に彼は一切返事を書いてない。太田氏もそうです。どちらが嘘をついているかは明らかです。
では、どうしてこんなことになったか、という思考順序になっていくんですね。今頃「すり替え、入れ替えなど、個人的な犯罪的行為をできるだけ考えないで、誰でも納得のいく説明はないものか?」と考えているんですか?
(アルイミオウジ氏の最近の動向2)①SNPとSNVの違いをどう理解しているのか説明してもらいたい。
②129の特異的SNPとして登録されている箇所の数と、B6の特異的SNPとして登録されている箇所の数を教えて貰いたい。
③Ts.Marker 氏はそれぞれ1000万か所程度だが、理研が調べた箇所は129とB6で異なっている300万か所と言ってるが間違いないか。
>>
------ 追記 6/30 -----
マウスのSNPsは1000万といわれていますが、調査報告ではB6と129で違う300万を使用している。④近交系マウスの定義は最低でもホモ率98%で、ヘテロ残存率1.3%ですから、30億dpに対して4000万弱のヘテロアレルがある。②はこの4000万アレルの中にはないはずだ。(あったらSNPs判定はできない。)アルイミオウジ氏の言うヘテロSNPsとは何を意味しているのか、答えてもらいたい。
>>
*www.anim.med.kyoto-u.ac.jp/Kuramoto/contents/ExpAnimGenet_1_inbred.pdf
⑤公開データ登録されている幹細胞のF1背景分に関して、129/Sv-X1にCAGがあるかないかのアルイミオウジ氏解析結果を教えて貰いたい。
(学さんの誤解5)
さて、ここまで説明後に初めて本来の話に戻れる。学さんの主張です。
>>
近交系は、クローンマウスではないので、98%で一致しても、残りの2%で一致しなければ、減数分裂で配偶子に多様性が生まれます。今300万箇所のSNPsの話をしている。ホモになっている場所だけの300万か所です。アルイミオウジ氏の話でも書きましたが、ヘテロのSNPを調べたらこのSNPs解析自体が意味をなさないでしょうよ。父母どちらもホモにそろっているからF1を造った時すべてヘテロになる。色分けだと緑になる。これが判定基準です。緑でなく青だとかピンクになっている部分があったら、それは親のSNPがヘテロだったということです。つまり、ホモであるべき場所がヘテロになっているから若山さんの維持繁殖させているマウスコロニーの近交系が崩れているということが分かる。本来ホモだという前提です。原理的にヘテロもあるのだとしたら、つまり登録されている特異的SNPがヘテロであり得るのだとしたらそもそもこの解析は何を識別していることになるでしょうか。根本的な無理解です。
自分の維持しているマウスコロニーにコンタミが無いかどうかを調べてくれる商売があるんですよ。登録されているデータがヘテロもあり得るんだったら調べようがないでしょ。登録されている特異的SNPsデータは全部ホモだというのが私の理解です。コンタミがあるとその場所がヘテロになる。
緑であるべきところがピンクになっているのは129にB6がコンタミしている。緑であるべきところが青になっているのはB6に129がコンタミしている。つまり、ピンクは129のコンタミを、ブルーはB6のコンタミを意味しているんです。区別が必要です。
129には以下の可能性がある。
a.市販の129/Sv-X1 GFP無し
b.市販の129/Sv-ter GFP無し
c.若山さんの自家繁殖させている129/Sv-X1 GFP無し
d.若山さんの自家繁殖させている129/Sv-ter GFP無し
e.若山さんの自家繁殖させている129/Sv-X1 CAG-GFP(ホモ)
B6には以下の可能性がある。
a.市販のB6 GFP無し
b.市販の(理研の)GOFマウス
c.若山さんの自家繁殖させているB6 GFP無し
d.若山さんの自家繁殖させているGOFマウス
e.若山さんの自家繁殖させているB6 CAG-GFP(ホモ)
f.大阪大岡部氏より移譲のB6-Acr-CAG-GFP(ホモ)
g.若山さんの自家繁殖させている岡部氏より移譲のB6-Acr-CAG-GFP(ホモ)
ピンクはa,c,dの可能性があるが、a.ではないはず。市販のものにこんなにコンタミの有ることは考えにくい。ntESG1,G2はb.dのどちらかだが、ほとんどコンタミは無い。
ブルーは全部gです。
もう一度貼り付けましょう。
ピンクはすべてに共通しているが6番染色体のみが「僕のマウス」グループと違っている。ここは129 cag-GFP mouseは緑になっている部分です。
それに対してブルーは元の岡部マウスのデータが無いが、「僕のマウス」グループ以外は全部自家繁殖の岡部マウスです。ここにコンタミのある部分が青で出ている。その出方に3種類ある。
①FLS3,129/GFP ES.CTS1,FES1
②FES2
③ntESG1、G2
なぜ3種類になるのか。まず最初に作られたのは③です。太田さんが岡部研から持ってきたときの岡部マウスで作ったもの。11番と18番にコンタミがある。
②は11番に③とは別のコンタミがある。ただし、この129はterではありません。なぜなら129 cag-GFP mouseのピンクパターンが共通しているからです。
①は6,11,12番に共通のコンタミが見られる。しかも11番は②と共通している。
以下が、学さんが言いたいことですね。我々の長年の関心事でもある。
>>
もっとも、調査チームが、FES1 FES2問題を論じているのは、ここではないけどね。
調査チームは、幹細胞成立後に起きた塩基変異が焦点だ。
何度も出てくるが、桂報告書の以下の6頁部分が、FES1 FES2問題の鍵となると思う。今回 4 種の幹細胞には、第 6、第 11、第 12 染色体上に 129 に特徴的なクラスターが、また第 17、第 18、第 19 染色体等に C57BL/6 のクラスターが 認められることから、TaqMan PCR によって観察された 129 ホモの SNPs はこれら幹細胞 の作製に使用したマウスに存在した遺伝的背景の不均一性によるものと結論づけた。 ES 細胞 FES1 と FES2 でのみ異なる SNPs に関して、両者の遺伝的背景の相違によると 判断された上記第 6、第 11、第 12 染色体の SNPs クラスターを除外し、残った 1,290SNPs を用いて比較を行うと、STAP 幹細胞 FLS3、FI 幹細胞 CTS1、および、ES 細胞 129/GFP ES は同一細胞株といって良い程の高い類似性を示すことが判明した。従って、STAP 幹細胞 FLS3、FI 幹細胞 CTS1、および 129/GFP ES は同一の細胞由来であり、ES 細胞 FES1 と同 一、あるいはそれから派生した株の可能性が高い、と結論づけた。 FES1 FES2の作製時、親マウスがどのような状態であったかはわからない。
但し、同じ受精卵由来でないことはわかるが、両者のSNPが離れている原因が謎になっている。学さんの疑義は今述べた我々の疑義そのものですね。
そして学さんはその原因を以下のように考えるのだけれども、日本語の助詞の使い方が間違っていてとても誤解をうけやすいのと、もう一つは推測そのものに間違いがあってややこしい。韓国系の人は助詞の間違いは少ない。シナ系の人は助詞の使い方が難しくてよく間違えますね。
>>
それぞれ、129もB6の両マウスは、マウス交雑の結果ヘテロであることから、F1において、129/B6とならずに、129/129、B6/B6の組み合わせができうる。
2003年に若山研究室に入導入されたアクロシン入りB6マウスは、FES1 FES2の親となったが、その遺伝子の詳細は公表されておらず、やはり129マウスのコンタミは起きていた。
つまり、FES1 FES2のSNPの様相は、ヘテロな親から引き継いだものと、ES作成後に生じてきた1塩基変異を合わせて考える必要がある。学さんの今までの考えを理解すると、上の文章は以下のように直せる。
>>
それぞれ、129も
(→と)B6の両マウスは、マウス交雑
(→混入)の結果
(特異的SNPsが一部)ヘテロであることから、F1において、129/B6とならずに、129/129、B6/B6の組み合わせができうる。
2003年に若山研究室に入導入されたアクロシン入りB6マウスは、FES1 FES2の親となったが、その遺伝子の詳細は公表されておらず、やはり129マウスのコンタミは起きていた。
つまり、FES1 FES2のSNPの
(異なった)様相は、ヘテロな親から引き継いだものと、ES作成後に生じてきた1塩基変異を合わせて考える必要がある。
ここでも「も」の使い方がおかしいですね。学さんはシナ系の一世のとてもよく日本語のできる方ということでしょうかね。帰化人の2世で日本の学校に通った子はこういう間違いは絶対にしません。周りの子供たちから矯正されてしまいます。日本人がこのような助詞の使い方の間違いを絶対にしないのも同じ理由です。
でも、外国人、もしくは帰化1世の人だと分かれば簡単に理解できます。我々は日本語はプロですからね。事情が分かれば推測が可能になる。彼女が言ってることは我々の言ってることと全く同じです。
問題は彼女の推測自体が正しいかということです。「ES作成後に生じてきた1塩基変異を合わせて考える必要がある。」ということですが、上の図を見て、そんな推測ができますか。ntESG1G2にあった青はどうして他の細胞では消えてしまっているのですか。これは元のマウスが違うとしか思えませんね。というよりも、この実験が最初ですから、他の岡部マウスからこの時の青が消えた理由が解明されないといけない。加わったものより消えた方が問題でしょう。
岡部マウスは太田さんが若山ラボに来たことが契機で大阪大から移譲された。太田さんはその岡部B6マウスを使って、ntESG1,G2とFES1,2を作っている。
2005/1/20に凍結し、2007年にもう一度解凍して使用し再度2007/8/3に凍結したntESG1,G2にあったブルー部分のヘテロSNPsが、2005/12/7に凍結したFES1,2からは消えている。(Ooboeさん情報)途中ですが、今の考察と関連しますから、学さんのところに書き込まれたOoboeさんからの情報をここで確認しておきましょう。「Ooboeさんとパートナー氏の資料館」にもアップしています。
>>
現在、FES1、とFES2の調査用のES細胞サンプルの考察が続いておられますが、パートナーが入手しているそのサンプルについての情報をお伝え致します。参考にして下さいませ。
2019/12/04 URL 編集
Ooboe
前コメント、名前入力わすれました。ごめんなさい。
FES1などについてのパートナー入手情報
理研広報は、調査サンプルを受領し、解析を担当した、CDB非対称細胞分裂研の研究者に直々に確認され、その研究者はFES1とntESG1は山梨大、若山教授から取り寄せました。
FES2とntESG2は山梨、若山教授とは別のところから取り寄せました。
これと同じ内容は、情報公開室が確認しています。
今晩、続きをお伝え致します。
2019/12/04 URL 編集
Ooboe
続きです。
前記にありますように、理研広報や調査委員会事務局に調査用サンプルの取り寄せについて報告された、BCA論文の筆頭著者でもあった研究者は、BCA論文中では、大田氏からFES1、FES2、ntESG1、ntESG2、を取り寄せたことになる記述をしています。
世界に発信されたBCA論文と、食い違う内容報告を、理研広報など、理研組織内では共有確認しているわけですが、なにより取り寄せ依頼し、受領し、解析したご当人がこの食い違いについての諸々の事実を把握していると思われます。
2019/12/04 URL 編集
Ooboe
さて、調査用ESサンプルを取り寄せ依頼、受領、解析をされた、CDB研究者はなにゆえに、理研にとっての公式発表を担当する広報室や、法令遵守担当のコンプライアンス室などの対し、組織内部にだけに共有される情報を伝え、対外発表のBCA論文や、調査報告書とは食い違う報告をしたのでしょうか?
もし、広報などに伝えた、内容通りならFES1とntESG1は、山梨大で用意された調査用サンプルということになります。
そして、FES2と、ntESG2は、別のところということですから。FES1とFES2は、出所が違う訳であり、細胞作成された、場所も同じでない別々に作られた、ことになります。又は、株分けでしょうか?
いずれにしろ、FES1は、京都大、大田氏から、取り寄せたのではなく、山梨大、若山研から取り寄せたと、BCA筆頭著者は、理研内部組織に伝えていたこの事実は、客観的公的事実であることには、変りがございません。
2019/12/05 URL 編集
Ooboe
パートナーは、FES1などのサンプル受領についての、MTA契約文書の開示請求をしましたが、情報公開室は、解析担当研に確認した結果を、パートナー宛に回答書を送付しています。その記述の中に、解析担当研究者が若山教授に直接依頼したもので、MTAや取り寄せ手続きの公的文書等は交わしていません、とありました。
6月30日理研は機関として、コンプライアンス室が予備調査を開始するとの発表をしていますが、しかしFES1等の調査取り寄せに予備調査事務局は、関与していませんでした。
この事案は普通の研究者間でのサンプル提供、受領手続きではありません。機関として、公的に取り寄せるべき事案であった訳ですから、関与していれば公的事務局として、文書交付は為されたはずでした。
解析調査をする側は、サンプルを取り寄せの依頼するのですから、取り寄せ宅配経費は、依頼側が支払うはずです。しかし解析担当研は会計処理をされてませんし、又、山梨大学、若山研の会計処理も為されてませんので、若山氏が自費でFES1の宅配経費負担をして理研解析担当研究者に送付していたことになります。
ということから、依頼したのは、逆に若山氏側であった。というのが整合的に帰結されてくる訳です。
以上
FES1等については不明瞭な事実が更に重なって存在していますが、省きます。
いずれにいたしましても、真正な調査サンプルとしての資格が無いに等しいのが、 FES1等サンプルでありました。
2019/12/05 URL 編集ほぼすべて既知情報ですが、新しい情報の一つは直接やり取りした人がDaijiro Konnoさんだったということですね。でも指示は責任著者の松崎氏から出ますからね。
「BCA論文の筆頭著者でもあった研究者は、BCA論文中では、大田氏からFES1、FES2、ntESG1、ntESG2、を取り寄せたことになる記述をしています。」の部分、事実であるかどうかは別として、そもそも太田氏が出発点だということになっているから、訴訟法的な厳密な確認はしないで、そう書いたという言い訳はできます。変だと分かっていたろうがと責めることはできますけどね。
「FES1等の調査取り寄せに予備調査事務局は、関与していませんでした。」というのは重大です。ガヴァナンスが無いんですね。
ところで、今ため息ブログ氏がこの件に関して以下のように書いている。
>>
近交系に混入があってもはや近交系ではなくなり遺伝背景が均一でない、これでFES1とFES2のSNPsの違いを説明できると言っているんでしょ。それだけでは説明できないと学さんも言っていて、我々も今、ここでそれを考えているところですね。ため息氏は全然専門家でないし、また、素人であってもこの件に関してよく調べている人でもないんで、こんな人と議論していても意味無いでしょうと言ってるんですけどね。
ため息氏のブログを読んでいたら目にとまったのでコメントしておきましょうかね。同じように誤解している人は多そうです。
>>
今さら何を言っているのでしょうね。FLSもAC129と同じようにCAG-GFP遺伝子の挿入部位を読んだのです。しかし、アクロシンプロモーターが付いたGFP遺伝子とCAG-GFP遺伝子が連結していたため、本来読むべき配列ではなく、通常15番染色体にあるアクロシンプロモーターの配列を読んでしまい、「15番」と誤認したのです。15番にはCAGは無いと言ってるのに分からないんですね。アクロシンプロモーターを含むプライマーで探したから15番の内在性アクロシンプロモーターを誤認した。CAGのプロモーターを探していたら15番にはCAGはないから誤認するわけないでしょ。
この人はもと数字君とコンビの岡目何とか君じゃなかったかな。
何度でも貼り付けましょう。
右はCAGプロモーターを含むプライマーで挟んだ。
左はアクロシンプロモーターとCAGプロモーターの両方を含むプライマーで挟んだ。
左のサンプルを右のプライマーで挟んだら15番にはCAGプロモーターは無いから検出できない筈だよと言ってる。アクロシンプロモーターとCAGプロモーターの両方を含むプライマーで挟んだ時にのみ誤認があり得る。しかし、この場合左の18番に無いとされている結果もアクロシンプロモーターが無いだけという可能性があると言ってる。
そもそも、右はCAGプロモーターしか含んでいないプライマーなのは15番に誤認がないのでわかる。ではどうして左はアクロシンプロモーターを含むプライマーで探したのかね。分かってたんでしょ。アクロシンを3番に発見してもらいたかったけど15番と間違えたので遠藤様が登場したということでしょ。
放医研のお友達にはどういうゲノムウォークをしたから間違えたのだとちゃんとした説明をして欲しいね。無料だから間違えたのかね。この人はタカラバイオでは商売にならないから雇われないだろうな。こんな間違いしてたら、損害賠償請求されてしまう。小保方さんの弁護士は何をしてたのかね。それとも放医研の人は無関係だったのかな。
(学さんの誤解6)ここ2回の記事で学さんは立ち直ったようです。もう彼らの相手はせずに、問題そのものを相手にするということのようです。直近の最後の記事に関しては私は完全同意です。そういう姿勢であっちを相手にしないと決めてくれたら私はそこに戻って質問してもいいですね。私は学さんの知識に関しては疑ってない。特にTCRについて教えられたことと、丹羽さんのGFPはクレだと私の勘違いを注意してくれた。忘れてませんよ。
時々刺激して無いと寝込んじゃうという意味では他の人がやってくれますよ。私なんか彼らが工作を続けていてくれているから何度でも繰り返し、若山さんが犯人だと繰り返せるわけです。彼らが続ける限り永遠に若山さんが犯人だと書き続ける。どうしてかというと彼らがいつまでも嘘の工作をし続けているからです。彼らが永遠に黙り込んじゃったらどうしますか。誰も何も言ってないのに、一人で若山さんが犯人だと同じ事を何度も繰り返さなくてはならなくなる。そうなると読者はそれはもう聞いたよと読まれなくなってしまう。雑談コーナーは今黙ってしまった。気づかれたか。はは。
(学さんが相手しなくていいように)>>
一ヶ月前の質問です。まだ生きています。答えて頂戴。
1) 学とみ子説「T細胞はキメラに寄与できない」のだから、TCR再構成を使う実験の提案者(西川氏)やそれを受けた笹井氏等は免疫学・発生学・細胞学に無知な方だったの?
2) FES1、FES2についてとは何を問題にしている?
3) 桂報告書のどこにES説では成り立たないことが書いてあるの?
4) 「科学的事実は、その時から、全く変化しました。」→ 5年経過してどんな科学的事実が変化したの?
5) 「これだけの事実が出てきて」→ 5年経過してどんな事実が出てきたの?
6) 「桂報告書に、若山研究室の見取り図を載せた人と同じレベルです。」→ どんなレベル?
7) 「遺伝子状態」て何?どのような状態のこと?
8) 「遺伝子が欠失すると多能性が発揮される」→ 具体的なそのような例を示して頂戴。核移植とiPSは遺伝子欠失ではないよ、そして実験的な多能性の発揮はこの2つしかないのでは?
9) 女王様とフィンガーボールのエピソードは笑い話だったの?
10) 幹細胞作成時には細胞をソートしていないと思うのですが、ソートしているとはどういうことでしょ?1) 学とみ子説「T細胞はキメラに寄与できない」のだから、TCR再構成を使う実験の提案者(西川氏)やそれを受けた笹井氏等は免疫学・発生学・細胞学に無知な方だったの?
学さんは論文どうりのSTAP細胞(小保方細胞ではない)があるという立場で考えておられるので、その推測の中でT細胞は選別されて生き残ってない筈だと考えたんです。白血球の他の細胞が生き残ったのではないかと。
2) FES1、FES2についてとは何を問題にしている?
私も今問題にしていますが、私の場合は細胞が入れ替えられているという基本的視点からの問題意識です。言うまでもありませんが細胞の入れ替えが無いのなら犯人は小保方さんになります。特にGLSについては動きませんね。ntESG1G2は論文、ラベルと中身が違っている。細胞の入れ替えがあったことを示唆している。Ooboeさんたちの追っている問題です。学さんは若山さんが犯人だということを考えていないので、なかなかすべての情報要素を整合的に整理できないでいる段階だとみています。
3) 桂報告書のどこにES説では成り立たないことが書いてあるの?
そんなことは書いてない。既存ESだと言ってます。ただ、キメラの捏造を断定していない。それは私に言わせると政治的配慮なんですが、学さんは調査チーム内、或は理研内での対立を想像している、特に竹市さんは15番にGFPのあるマウスを調査中と報告している。理研には小保方さんを雇って問題を拡大した責任に関して各所に配慮があるという事情もある。そもそも大した問題でなかったものです。
4) 「科学的事実は、その時から、全く変化しました。」→ 5年経過してどんな科学的事実が変化したの?
5) 「これだけの事実が出てきて」→ 5年経過してどんな事実が出てきたの?
桂報告書記者会見の後に、より詳しいBCA報告がでて、ワトソン氏はそれを知らずに私と議論して、最後にブレンデルのピアノに関して喧嘩別れしましたが、あれはTs.Marker氏によるとDORAさんだったらしい。はは。その後再現検証実験についての丹羽さんの報告と相沢さんの報告がでた。米国でも再現実験の報告が出た。7月頃にはキンガ・ヴォイニーツの論文がでてティシュー論文が批判的に引用された。それといろんな人が既存の科学的事実から演繹した知見も増えてきているというような意味でしょう。
6) 「桂報告書に、若山研究室の見取り図を載せた人と同じレベルです。」→ どんなレベル?
アホでしょ。何も意味してない。
7) 「遺伝子状態」て何?どのような状態のこと?
これはこの言葉の使われた状態を知らないので分からない。
8) 「遺伝子が欠失すると多能性が発揮される」→ 具体的なそのような例を示して頂戴。核移植とiPSは遺伝子欠失ではないよ、そして実験的な多能性の発揮はこの2つしかないのでは?
2つ以外にミューズ細胞とムーさんの細胞があるかな。小保方細胞が何なのかは研究中ですね。今報告は止まってるが。ただ、「遺伝子が欠失すると多能性が発揮される」というのはどういう文脈の中で使われたのか、多能性獲得というのはリプログラムされることなので、欠失すると遺伝子情報が不完全になるという意味なので、遺伝子欠失と多能性発揮を結び付けた研究は誰もまだしてないのではないか。なにしろ、学さんはSTAP細胞有りの前提で考え続けられている過程での試行錯誤ではないでしょうかね。
9) 女王様とフィンガーボールのエピソードは笑い話だったの?
暗喩というのは何を裏に隠すかということなので、意図は本人でないと分からないし、分からないときに無理にわかる必要もない。何事かをあからさまにしたいときには比喩は使わない。
10) 幹細胞作成時には細胞をソートしていないと思うのですが、ソートしているとはどういうことでしょ?
以前から気になってましたが、木星リストのFLBはソートされていると書かれているんです。まず事実を確認して後に、このソートは何かということになる。我々は無論ntESをキメラ胚に入れた後にそれを幹細胞としたわけですが、ここにはリシピエントのインナーセルマスが残るので、GFPでそれをソートしたと考えている。テラトーマの体細胞はソート前の幹細胞を若山さんが上から注射したからリシピエントのインナーセルマス由来のES細胞がテラトーマ化したとみている。これは2Nでも4Nでもソートされている。4Nでも胎生致死以前の段階なのでまだ死んでないと見られている。和モガさんも考察しているところです。
- 2019/11/25(月) 09:40:06|
- AC129
-
-
| コメント:11
(本題に戻る)犯人は既に若山さんだと分かっている。アクロシン発見に至る経緯は白々しい小芝居だというのは理の当然ということになる。
①自分は「僕のマウス」を渡したのだという嘘が起点になる。証拠は何もない。実験ノートの記載すらないことになっている。
②FLSを作成している頃には小保方さんの後の論文記載によればGFPはB6側にのみ入っている認識で、それに対する共同著者である若山さんの否定も無い。
③4月にヴァカンティが米国特許仮申請をした。
④その後に当時結果の出たジャームライントランスミッション実験で、小保方さんに対して、GFPが半分にしか来なかったという情報を刷り込んだ。そもそもヘテロなら半分にしか来ないのが当たり前だが、ここであたかもホモで渡していたつもりだったんだというアリバイを作っている。実際には半分にしか来なかったのが事実なので、ヘテロだったということで、小保方さんはこの時にサンプルに+/-表示した。そもそもキメラ子以前にその親は全部オスだったのに実に白々しいことをしていると今となってはわかる。
⑤その後「僕のマウス」でES細胞を作って小保方さんに渡した。
⑥AC129の実験の話はその後の出来事である。
私の手法ははっきりしていて、若山さんが犯人であるから、何をしたかと推測していく作業です。若山さんが犯人であるのだからこうしたはずだと推測して、実際にその痕跡が無いかを確認していく。仮に私の出発点が間違っていたら必ず深刻な矛盾が出るということです。今のところ出てない。私としては難問にぶつかるたびにどうしてこんなにいつも考えた末には問いが解けてしまうのだろうかと驚く。逆の立場で考えていた初期には経験しなかったことですね。多分出発点が正しかったのだろうと推測している。
Ooboe さんは私が考え続けている意味を誤解されているかもしれませんね。若山さんが犯人だという確信は何も動いていません。私にとってはもう結論は出てしまっているんです。でも、だからと言って、彼の行ったことの全てがわかったわけではない。分かってないことの中で最大の難問がAC129なんです。これは基本三誌論文とは無関係で、TCR再構成検証、胎盤蛍光問題、ローザ、そして8月のGRASへの試料提出へと繋がっている問題で、小保方さんは後のレター論文に書いているようにAC129でキメラが作られたと聞いていること、そして公共データに登録されている諸細胞が基本F1であって、129ではないということとの間に大きな矛盾があって解明を待っている。だから考え続けているだけです。
L氏はこの分野の専門家らしいので私の専門知識の足りないところを補ってくれるのではないかと思って、Ooboeさんは議論してもらいたがっておられる。その議論を聞くことによってご自分の理解も進むと期待されている。
まず大間違いの認識を改めていただきたい。このSTAP問題をネットで扱っている人々に本物の専門家は一人も居ません。相沢さんは事件に関してあなた方とは話さないとおっしゃりませんでしたか。丹羽さんがこの件に関してあれから何か語られましたか。清成さんも何もおっしゃってませんね。私と例えば丹羽さんと議論をしたらOoboeさんはその議論を通してご自分の理解が進むと期待されている。丹羽さんでなくてもいい。本物の研究者とど素人代表の私が対話して、専門家の意見を聞き出してほしいと。大間違いなんです。
本物の専門家が私の質問に答えてくれるなんて思っていることが大間違いです。専門家が話せる位なら私が質問しなくても既に本が出てますよ。彼らは語らないと決めているんです。語っているのは有象無象です。Lさんもそうです。英国に何しに行ってるんですか。明治の学生は国家のために勉強した。ホームシックにかかって時々日本のブログに書き込んで寂しさを紛らわしているような暇なんてあったら勉強してますよ。
こういう状況で誰と対話しても得るものはありません。残念ですが、官僚制度の弊害が及んでいて、ラブオンザビーチというのは社会問題ですからね。STAP事件問題ではありません。ときどき羽蟻が飛んでるのをみかけるようになったら既に社会の基本構造に白アリがたかっていると見ないといけませんよね。
わたしは常時チェックしているんですよ。
>>
5558. 澪標
2019年11月12日 13:46
Lさん
一言居士さんの「長い記事」とは、「AC129を巡る問題、1~8<以下Ac129.Xと表記>」だと思いますので、これを前提にコメントします。
一言居士さんによるAC129.X論考は、学術的論考の規矩をはるかに逸脱したものと存じます。一例を挙げると
❶AC129.6ではSTAP様塊のキメラ実験においての、実験を実施した清成さんの意図的ともいえる沈黙
❷AC129.9ではChiP-seq lysateの存在有無を巡っての、伊藤さんによる意図的対処
それぞれの存在に言及していますが、その存在”立証法”は極めてレトリック(排中律の存立保証のない状況に置いての排中律の多用等)に満ちたものであり、プロパガンダとしか言いようがありません。
プロパガンダであろうとなかろうと、一言居士さんご本人の主張(便所の落書き)に沿うもので限りにおいては、等閑に附せばよいことです。
しかし第三者が別の場で、一言居士さんの所論について考察対象として、広く意見を求めるとなると別問題です。
Lさんご自身が、一言居士さんの所論、立論方法、措辞の置き方について総括的評価を行って後にこそ、個別問題への言及・他者の意見の募集が可能と愚考いたします。
<続く>
5559. 澪標
2019年11月12日 13:50
リゴリスティックにすぎるとお考えかもしれませんが、当ブログ、したらば掲示板、現在の自己ブログにおける一言居士さんの言説を振り返れば当然の帰結と存じます。
くり返しになりますが、一言居士さんの言説でまず問題となるのは「主張の当否」ではありません。「主張がDue Processに則ったものであるか」です。
敢えて申し上げます。興味の赴くままに話題を拾い上げ、言葉を発するのは幼児の伎と存じます。
蛇足ですが、私の一言居士さんの発言への評価は
和田家文書(東日流外三郡誌)についての古田武彦博士の論考と準同型の、”アプリオリに措定された結論<博論記載のC.Vacanti留学時代の一連の実験は真実>”から倒立的に構築されたポレミックなファンタジーです。
<博論記載のC.Vacanti留学時代の一連の実験は真実>について、色々論考されていますが、正典(「あの日」&「博論」&「Tissue Engineering論文」)と外典(「小保方晴子日記」、「不服申立書」、「不服申し立て時の記者会見」、「STAP発表時の記者会見」、「STAP HOPE PAGE」)の間に存在する矛盾点には一切言及していません。ここに書かれていることはすべて間違いでスピン記事に過ぎない。彼は正しくスピン屋としての仕事をしているんです。でもL氏に関して「敢えて申し上げます。興味の赴くままに話題を拾い上げ、言葉を発するのは幼児の伎と存じます。」という批判部分は単なるストレートボールです。私のL氏への評価は以前からこれです。若くて思考が浅い。多分語り合っていると彼の専門知識は私の参考になることはあるでしょうね。でもそれ以前に思考の深度が噛み合わないと思いますね。相手が何を考えているかに思い至らない。これは持って生まれた性格もあるんですよ。物を考える仕事に向いてないかもしれませんね。違う分野の方がいいかもしれない。
上の引用に対するLさんの理解は以下です。最後の部分ですね。澪標なる人物は私の論に対してスピンをかけているんですけど、Lさんが理解したようには言ってませんね。L氏はまず相手の思考を理解するという基本的な作業ができない性分です。これはスピン屋として意図的に誤解を作っているのではありません。
5580. L
2019年11月14日 04:45
5560. 体内時計さん
>Lさんが仰る「意図」とはなんですか?
「意図」が何か不明と言ってますから、分からないという事です。モンキーさんのコメントにもありますが、使っていないRosa GFP knock-inを筆頭著者が自力で持ち出すとは考えにくいですよね。一言さんの記事を見て、誰かから、何らかの意図で、このマウスについてのインプットを得た筆頭著者が、このマウスを使ったと思い違いして書いてしまったと推測しましたが、モンキーさんの推測も有り得る話だと思いますよ。いずれにせよ、幾つも有り得る可能性のうちの一つでしかないですから。信頼できる情報が全然無いので、可能性を絞れませんから、疑問点としては残りますよね。
モンキーさんにしては珍しいと思うのですが、引用されたコメントにはたくさん推測が入っていて、簡単に納得はできないと思いますよ。ご自身の考えにフィットするコメントについては、推測であっても「納得」し、合わない推測について反論する、というのはバイアスですね。
一言さんのご意見は、澪標さんがおっしゃるように、彼の仮説に合うものをつなぎ合わせたもので、バイアスまみれですが、その中に一部でも真っ当な考察があれば、私は参考にしますよ(今回は読み込む暇がないですが)。体内さんのご意見についても同様で、モンキーさんの推測は、有り得るストーリーの一つだと思いますので、参考にさせて頂きます。(が、即納得するわけではありませんので、ご理解を)「一言さんのご意見は、澪標さんがおっしゃるように、彼の仮説に合うものをつなぎ合わせたもので、バイアスまみれですが、」という理解が既に間違ってる。澪標氏は「当ブログ、したらば掲示板、現在の自己ブログにおける一言居士さんの言説を振り返れば当然の帰結」と書いている。そこから出発してわたしがデューを守ってないという具合にスピンを掛けていくんです。L氏ではスピン屋にもなれないでしょうね。
澪標なる人物は私の方法論が筋が通っているということを認めているんです。それが「当然の帰結」という言葉に現れてくる。私が「仮説に合うものをつなぎ合わせたもので」矛盾なく事件の結末を説明できているということに関して否定していない。私の方法論を否定するのなら、私の構築したストーリーに関して個別具体的な矛盾を指摘しないといけない。
逆に私は桂報告書のストーリーに関して個別具体的矛盾をたくさん指摘している。ここの「桂報告書疑義」に網羅している。
彼は私の諸論考の矛盾指摘ができないので、科学的方法論にまず法学的概念であるデューを捏造して、スピンの手法としている。指に唾をつけた瞬間ですね。論考はただ合理的で内部諸矛盾がなければそれでいいんです。科学の場合には物的実証が出来たらそれでいい。デュープロセスなんてありゃしない。
実証は素人側ではできないので、結局は理研チームの行った結果に関しては基本それをよりどころとするよりないんですね。ど素人は実験できないからね。専門家でも予算無しではできはしない。怪しい奴らだけど、科学者としての誇りくらいは持ってるだろうという仮定で信じるということですね。本当のことは隠すにしても提出しているデータが嘘ということでは自ら耐えられないであろうと。
まあ、彼らが例えば一つだけ。ntESG1のラベル記載と中身が違っていたということに関して何か合理的な説明ができたか、或は説明しようとしたかということを考えたらわかる。彼らが行わねばならないのは自分たちのストーリーである小保方さんのESコンタミという趣旨に対しての疑義に解を与えることです。で、相手の説を批判するときには言ってることが矛盾していると具体的に指摘すればいいんです。彼らはどちらもやってない。だから鉄さんにクソがと言われるんでしょうね。
(基本事実)
キメラは出来ている。
①論文通りにキメラができた。
②小保方さんが若山さんを騙して既存ESを渡したことによってキメラができた。
③若山さんが小保方細胞核を使ってntES化させたキメラをリクルートの都合で一時的に直接できたかのように語った。
④若山さんが誰も思いつかないような手法でキメラを作った。
⑤その他
Ooboe さんに申し上げておきますけど、①は無いんですよ。というのも再現実験してますよね。出来てない。論文発表するということは他人はともかくとして、論文の著者たちには簡単に再現できないといけないんです。条件が少しでも変わったらできないというのは、まだ解明途中なので、論文で公表されるべき段階ではありません。論文には何度でもキメラができたと書かれている。従って、いろんなことは考えられるにせよ、論文通りのSTAP細胞は無いという言辞は正しいんです。
①ならば論文通りにキメラは出来たのだが若山さんが参加しなかったから再現できなかったということ以外にありません。出来ている細胞を若山さんが隠したということです。若山さんが犯人だということですよね。なぜそんなことをしたのか考えないといけない。
②は桂報告書のストーリーです。矛盾指摘は嫌というほど行われていて、誰も解を発表してない。今パートナー氏だけが唯一行動されている。他は私も含めて話しているだけだ。
③これが私の説です。今のところ矛盾指摘はない。無論分かってない謎は多いので、ここが明らかになって結論が否定されるということはあり得ることでしょうね。でも今のところ何もわかってない。
④⑤はお手上げですね。
(私は何を考えているのか)L氏は学生なのではなくて研究所か何かに勤務されているようてすね。
>>
5603. L
2019年11月17日 06:58
一言さんは、細かい解釈の積み重ねで論理展開するので、きちんと読まないと意見できないんです。あまりに長いので、最後のところだけ読んで、ちょっとおかしいなと思うところだけコメントしました(最後のところはそれまでの本文とは完全に独立しているように見えたので)が、まさか2番のところで噛みつかれるとは思いもしなかったです。とりあえず1番と3番がおかしい事をお伝えしたかったのが本心です。本文は今の仕事がひと段落ついたら、よく読んだ上でコメントする事にします。
モンキーさんのコメントに納得できないのは、あまりに笹井先生に失礼な推測だと思うからです。笹井先生がそんないい加減な形でメソッドを書くとは考え難いです。なので、筆頭著者が書いた(あるいは筆頭著者からのインプットで笹井先生が書いた)と私は推測します。この場合は、まず、筆頭著者に誰かからRosaマウスについてのインプットがある必要があると思いませんか? だとすると、どのような意図でインプットがあったのか、疑問に思いませんか? 謎でしょ?
5604. L
2019年11月17日 07:37
あと、Oct4抜きのiPSの論文がちょっと話題になってますね。Oct4があるとリプログラミングの質が悪くなるとの事で、Oct4をマーカーにしたSTAPでは限界があったという事でしょう(仮に何らかの現象が筆頭著者のOct4-GFP陽性細胞に起きていたとしても)。
私の考察結果は一枚報告と一枚報告補記にある。ここは考察ではなく結論です。AC129を巡る問題は論考中ですから、分かりにくいのは当然で、本人が分からないから考えているわけです。ただ、考える前には必ず直感がある。それを述べておきましょう。
小保方さんのそもそもの論文の主旨はシンプルな主張です。細胞をもちゃもちゃ扱っていたらPCR検査でOct4発現する細胞をみつけ、それを培地誘導したら三胚葉に分化した。テラトーマはできなかったが、ヴァカンティ足場を使ったらテラトーマ様の分化があった。理研に来て、白血球を酸浴させたら、Oct4-GFPが大量に発現するようになり、PCRで確認したら、内在性Oct4遺伝子も発現していた。キメラを若山さんに作ってもらったらできなかった。だから米国に戻ってそのできなかった結果も含めて論文に纏めて発表しようとした。2011年11月のことです。
11次元氏はここまでの間に小保方さんが捏造していると言ってる。一番大きな要素は三胚葉分化させたことですね。ESを混ぜたんだよと言ってるのと同じです。
こんな論文はどこにでもあるでしょう。これが何かは今からの解明を待つ話ですよね。何か妙な発見があったよと言ってるだけだ。ここに小保方さんがES細胞をポトリと垂らして三胚様分化させて、発見だと騒いだのだと言ってる。面白い男ですね。
ヴァカンティ氏はこの時点(ティシュー論文発表時点)では特許申請していません。論文発表にとどまっている。小保方さんの博士号取得を待って自分のところで雇用し、この研究を続けさせてその成果を特許申請しようとしている。
ずっと後になって、笹井研で小保方さんはこの試験管内三胚葉分化実験を行って見せている。当然この時もES細胞を垂らしたということになりますね。ヴァカンティ研で見た仲間たち、東京女子医大で大和氏が見たもの、恐らくは若山研でも見せたはずの、そして笹井研では笹井さん自身が見たと言っている三胚葉分化実験はESだったのだということになる。これが11次元氏が主張していることですね。
11次元が指摘したティシュー論文の瑕疵はTCRのバンドの写真が捏造だというものです。この件に関してはヴァカンティ研から説明があって、手伝ってくれた仲間のミスだと小島氏が謝罪した。つまり、事実図の間違いはあったということです。
で、11次元氏はこれをどうするつもりなんでしょうね。ESコンタミだと言ったのは彼ですよ。この始末はどうなっているのか。
更に、再現実験で、これほど確実な検証チャンスがあったのに、三胚葉分化確認実験が行われたのか否かの理研発表が無い。これはキメラ作成なんかとは比較にならないくらい簡単に出来る検証です。私はやっていると思ってますよ。なぜならここで三胚葉分化しなかったら、小保方さんの捏造は確定するじゃないですか。事件は終わってしまいます。
ここで三胚葉分化した結果が出ていたらどうなるか。これってただティシュー論文、博論に捏造が無いということが証明されるだけだ。理研でのネイチャー論文が捏造であるか無いかとは無関係だ。しかし、11次元氏には落とし前をつけてもらわないといけませんよね。どれだけ騒ぎまくったか。この人が誰だったか、正体を暴くべきでしょうね。
三胚葉分化実験の結果に捏造がないのなら、早稲田大学は小保方さんの博士号を剥奪することはできません。理研で何があろうと関係ない。
でも、早稲田大学が博士号を剥奪した理由は小保方さんが自ら博士号を一旦返上して再試験を望んだので再試験の結果博士号を与えなかっただけです。ティシュー論文と博論に捏造が認められたということではありませんね。早稲田はそんな証明はしていないどころか、事前の調査で博士号剥奪は出来ないと結論しています。彼女は自分から返したので博士号を失ったんです。その時早稲田大学がどんな説得をしたのかは又別の事件ですね。再試験して再付与すると騙したんでしょ。何しろ草稿を提出した落ち度と、それを確認もしないで受け取った落ち度とをバターにしたということですよね。で、正しい本論文を出したけど今度は落第させたということです。私は詐欺に類した行為で大学は小保方さんの支払ってきた授業料は全額返還しないといけないんじゃないかと思いますね。
キメラはこの後にできたんです。若山さんは別の実験を始めてのだというのが私の仮説で、その仮説で事件を追っていくと何もかもがうまく整合していく。その結果は一枚報告とその補記に書いた。でもまだ分かってない問題がある。キメラができたにとどまらない、幹細胞化研究の実態です。それを今、AC129を巡る問題として考えている。
若山さんが犯人なんです。犯人は何をしていたのかと考えているんです。一旦自分の仮説を造ったら安易に立場を崩してはいけない。ちょっと難問にぶつかったら反対側に振れるようでは、逆にそちらでも難問にぶつかって、あっちとこっちと行き来する。とことん自分が間違ってると納得できるまで仮説を変えてはいけない。
放医研なんかに頼まないで、タカラバイオに金を払ってちゃんと調べてもらったら内在性アクロシンと間違えたなんてど素人みたいなことは言わないでしょうよ。批判するのであるならちゃんと私の思考計画に沿って批判助言してもらいたいね。私が何を追っているかが分からなければ批判もできないでしょう。遺伝子解析は高いお金を払えばちゃんとその道のプロがやってくれる仕組みがもうできている。機械そのものの値段が高いから減価償却費が高くなる。だから一件数十万という検査費用になる。でも数十万払ったらちゃんとやってくれるから商売として成り立っているんです。間違えて済む商売だと思ってるんですか。賠償問題になるよ。それくらい確度高くやれる技術があるんでしょ。放医研が何を間違えたって? その理由が理解できるから①の問題は解決済みだって? 無料だったからいい加減にやって間違えた挙句小保方さんの印象を悪くするのに役立てたって? 頭大丈夫か? 警察だったらわざと間違えたふりをしているのだという疑義から検討し始めるでしょうね。
手記を読んでないという時点で既にこの問題を論じる資格が無いんだよ。小保方さんが犯人なら、手記は犯人の書いた言い訳だ。警察は喜んで読むだろうね。書かれていることの裏を取ればいいだけだ。彼女が犯人であろうとなかろうと犯人だと疑われている人間が語っていることは事件のカギになる。光る精子で顕微授精してましたって書かれれば、ああ岡部マウスはいたんだよな。若山さんは常時維持繁殖させているんだよなと気づける。アクロシンが出たとなったら、まず自分の維持繁殖から疑うべきでしょ。どうして6年も前の太田ESが出て来るんだい。ほんとに「僕のマウス」を渡したのか。実験ノートに書かれてないじゃないか。何か証拠出せよということでしょうよ。遠藤氏が調べたらアクロシンだったって? 臭い芝居するなよ。
そういう風に疑っていくんだよ。そしてその自分の疑いが間違いだと証明されたら逆が証明されたことになるんだ。どちらから疑って行っても同じ結論に至れるのさ。情報がフルに出そろっていればね。ミッシングリングがあるときはいくつかのか可能性が残ることになる。僕は一つ示している。理研も一つ示したが内部矛盾を指摘されて否定された。L氏はどういう可能的解を示すのかな。そして僕の解に矛盾があるのならどう指摘するのかな。恐らく手記を読みもしないで何のストーリーも描けないと思うけどな。必要な情報で欠けているものがありすぎる。若山さんは彼女をリクルートしようとしたということを手記を読んでなければ知らないはずだ。裏返すとなぜ小保方さんが理研の客員で研究することになっていったのかを知らない。これだけでも既に致命的な知識不足がある。
(AC129に至る経緯)1.ヴァカンティによる米国特許仮申請
2.ジャームライントランスミッション結果が「ホモを渡していたのにヘテロだった」という小保方さんへの刷り込み情報
3.「僕のマウス」ESの作製(129/B6)
4.胎盤蛍光STAPキメラの発見(129/B6)
5.CTSの作製、胎盤蛍光CTSキメラの作製(129/B6)
6.「僕のマウス」TSの作製(129/B6)
7.ネイチャー誌のリジェクトを受けてのTCR再構成確認実験
8.幹細胞に関して小保方さんに直接やらせなかった
9.幹細胞とキメラTCR結果が若山さんの予期した結果で無かった可能性
10.AC129の実験(129ローザ)、キメラ作成
11.B6/129による何らかの実験(ntESG1、G2 ?)、公開データ登録情報(B6/129)
12.GRAS提出試料(129/B6)の実験?
13.GRAS提出STAP細胞(lysate)が「僕のマウス」ESになるようにした犯罪行為
14.AC129の中身を「僕のマウス」ESに入れ替えた犯罪行為
この時期にハーヴァード側と特許でもめている。そして10月の末に小保方さんがヴァカンティの許に帰ってしまう。その間に何が行われていたのか。この辺りの解明が求められている。
1.ヴァカンティによる米国特許仮申請ヴァカンティはティシュー論文の成果では特許申請していない。キメラができたと聞いて、そして小保方さんが最初のネイチャー投稿論文を書いた時期に合わせて特許の仮申請をしている。無論小保方さんが筆頭著者でヴァカンティが責任著者、そして若山さんが共同著者に名を連ねていたでしょうね。私の仮説ではこのキメラは小保方細胞核使用ntESからのキメラですが、彼女とヴァカンティはそれを知らない。2月頃に若山さんは小保方さんを山梨大の助手で勧誘している。前年の勧誘を入れると2度目の勧誘です。人事秘の解けるここまで彼女を引き留めるための便宜的嘘だったのだと推定している。ここで彼女が行かないとはっきり返事しなかったのが事件の遠因と考えている。無論、行くという返事はできません。彼女はこの時の現在ヴァカンティ研の所属ポスドクで、給料をハーヴァードからもらっている。
恐らく彼女はまだ論文が書かれていないのでヴァカンティとの約束が果たされていないと答えたでしょうね。10か月後結局彼女はヴァカンティ研に帰ろうとしたんです。つまり山梨に行くという明確な意思は無かったんです。それをずるずる引き延ばした結果になった。4月にネイチャー論文を提出した。約束は果たされた。でもリジェクトされた。ESコンタミだよ。なんて杜撰な研究室だとあっさり拒否された。
ヴァカンティ氏はこの論文をティシュー誌に掲載し、論文は出たのだから約束通り、小保方さんを解放してくれて、若山氏は望み通り小保方さんを山梨大に連れていける筈だった。
でも、ヴァカンティ氏はそうせずに米国特許仮申請して、小保方さんにはメジャー誌を目指しなさいと励ました。キメラができているのなら当然の判断ですね。
2.ジャームライントランスミッション結果が「ホモを渡していたのにヘテロだった」という小保方さんへの刷り込み情報若山さんは小保方細胞核は何物かだと思っています。キメラを作って見せてやったのはただ単に翌年に助手勧誘するための時間稼ぎのいたずら理由だけではない。なぜならば、もしそうなのなら、彼女が断った時には、何かの実験ミスでキメラができたようだといえば終わる話です。他所の誰かに頼んで確認してもらってくださいとやんわり縁を切ればいいだけで、ヴァカンティに対してもそんなにむつかしい言い訳でない。
小保方さんは行くということは言えないが、逆に自分の意思で行かないということは言えたんです。でも10か月後には帰ってしまったほど行かない可能性も自分の心の中に抱えたまま、行かないとは言わなかった。言ったら縁が切れて論文完成まで行けないかもしれないとは考えるでしょうね。
仮に彼女が桂報告書の主張するようにESコンタミさせた捏造犯人だったとしたら、捏造した結果は論文にならないとどういう意味でも評価になりませんから、論文までは書かないといけない。書いた結果は若山さんのところで研究を続けるのか、ヴァカンティのところで研究を続けるのか、いずれにしても再現はありませんよね。歴史的経緯としては理研に採用されて、特許申請し、自分は逆流性食道炎モデルマウスからSTAP現象を確認する実験計画書も提出しているが、出来はしませんね。どうせESだったんでしょ。そもそもGOFマウスを使ったテラトーマ実験で持ってる学生のntESを捏造に使わないということはありません。CAGのESを使ったら蛍光してしまう。こんな馬鹿な捏造なんてあり得ません。
若山さんが犯人です。若山さんは捏造なんてしていません。ただ、方便の嘘と、勘違いの実験をしていただけです。これだと人間性の自然からして理解可能なんですね。
若山さんはヴァカンティが特許申請した時万が一の時の言い訳として「僕のマウス」での実験だったという嘘の言い訳で逃げる計画を考えた。彼はそもそもヘテロマウスを使っていたので、大本のマウスがホモだということにしておけば実際がヘテロならESコンタミだったと言い逃れると考えたわけです。
でも、若山さんはすぐにそうしてヴァカンティをあきらめさせるという行動に出なかった。その原因が若山研で初めてOct4-GFPが大量に光るという現象が確認されたからです。キメラはできなかったが、若山さんはこれは何物かだと信じたんです。彼はntES化実験を本気で行っていて、まだ結果が出ていなかった。だからホモの話はすぐにはしなかったんです。
3.「僕のマウス」ESの作製(129/B6)3と4の前後は不明ですが、「僕のマウス」ESは細胞リストには2012/4/19が樹立開始日と書かれている。しかし、若山さんの持ち出しリストでは2012/5/25となっている。報告書は出来上がったのが5月の半ばと書いているんですね。4/19という具体的な日付は実験ノートに書かれているのだと思われる。ヴァカンティの特許仮申請は4/25です。事前に小保方さんから伝えられているでしょうね。彼はまずいことになったなと考えていたでしょう。ただ、論文は落ちるという自信があり、事実落ちた。これで終わりだと思っていたら、ヴァカンティは諦めなかった、当たり前ですね。若山さんがあれはESコンタミだったみたいと言わない限りはあきらめませんね。
或いはあれはntES化してみたんだと本当のことを言わない限りはヴァカンティは諦めない。でも後者は結果が出ていない。研究中だ。自分の研究なのだからヴァカンティに本当のことを言うつもりもない。若山さんは小保方さんをリクルートしたいという望みと、自分のntES化技術の応用としてのSTAP研究を山梨で行いたいという望みを同時達成したがっている。二兎を追っている。
4.胎盤蛍光STAPキメラの発見(129/B6)若山さんはキメラの胎盤が光っていると思った。欲目かもしれない。小保方さんに胎盤をいくつか渡した。つまりこの時、2月初頭の実験とは別に新たにSTAP細胞を作らせてSTAPキメラを作ったことになるが、既に検討したように、このキメラはFLSから作られたキメラではないかと思っている。ただし、STAPキメラだと言って胎盤を渡しているのだから、17日程度前には赤ちゃんマウスを渡していることになる。実際には使ってない。FLSの4Nキメラの胎盤だと思われる。小保方核使用ntESの胎盤が光ったということである。小保方さんに免染させた。
今となってはこれは小保方細胞核が原因ではなく、ntESの性質かもしれないとも考えられる。若山さんは当時未知であったGFPの漏れ出し蛍光に騙されて、小保方細胞核を移植したと思っていたが、その確率はとても低くて、単なるCD45細胞の核を移植しただけである可能性の方がはるかに高い。
5.CTSの作製、胎盤蛍光CTSキメラの作製(129/B6)手記によれば胎盤が光ったという話になったのは2012年の春です。ところが桃子本では時期は分からないとしている。情報源は理研若山研の中で、関係者と書いているので、若山さん本人か奥さんの可能性が高い。他のラボ仲間は何も言えない筈なので、まあ、奥さんだと思っていて間違いないでしょうね。
なぜ2012年の春だとちゃんと言わないのか。時期を明示して何が悪いのか。STAPキメラというのはSTAP細胞の維持培養が1週間程度しかできないので、この時のキメラと言ってしまうと小保方さんに渡した赤ちゃんマウスがいつのだと小保方さんに分かってしまう。
桃子本では胎盤が光っていると言い出したのは誰か分からないということになっていて、記者会見で若山さんも誰だったか忘れたと答えているが、手記では若山さんから光っているから免染確認してくれと渡されたことになっている。桃子本では小保方さんがみんなの前に胎盤を持ってきたところから始まっているが、胎盤を切り出したのは若山さんに決まっている。何も取材してないということがわかる。質問もせずに言われたままに情報を垂れ流しているだけだ。特ダネのためなので、生活のための犯罪ですね。こういうのを羽織ゴロツキという。ジャーナリストではない。
胎盤をいくつか受け取った後に4月には後のFI-SCの研究に入ったようだと書かれているので、時期が3月末か4月の始めと推定できる。2月の初頭からFLSの実験に入っている。1か月以上後のことなので必ず、FLSとは違う時期に赤ちゃんマウスを渡していることになる。
FI-SCはGOFマウスでは作られていないという証言になっているが、Oct4-GFP蛍光している写真が論文に添付されている。若山さんは作った記憶が無いと言っていて、作って無いと断言していない。作っていなかったら論文添付写真は捏造だということになる。誰が撮影したものかと問われなければならない。桂調査は何も報告していない。
どちらかが嘘をついているんです。犯人はどちらかで、小保方さんのESコンタミ捏造は無い。すると若山さんということになるんです。若山さんということになると、ではなぜという問題が生じる。このことについて考えているのはここだけで他にはない。今もない。みんなでだんまりを決めこんでいる。これが文科省の判断なんです。その板の裏を捲ると白アリが大量に出てくるからです。若山さんを守るためではない。はは。スパイが多いねえ。そろそろ摘発しないといけないんじゃないかな。国外追放だな。
6.「僕のマウス」TSの作製(129/B6)FI-SCは笹井さんがとても関心を抱いた現象ですが、あくまでもSTAP細胞のデリヴァティヴズとしての関心で、事実は単にntESの性質であった可能性が高い。ntESに胎盤異常が多いという原因との絡みで発生している現象ではないかと今となっては推測される。これはこれで若山さんの研究としてはとても重要な課題になりそうですね。小保方さんの細胞が何であるかとともに、それぞれ別途研究課題となるものだったでしょうが、どちらも今やできなくなってしまったのではないかな。
小保方さんの細胞は論文通りに三胚葉分化しなかったらただの捏造論文です。私は最初小保方さん犯人仮定で追っていました。理研でのキメラが小保方さんの捏造だったら三胚様分化もESによる捏造に決まっていると直感しましたから、早稲田の調査結果を読んでからティシュー論文を求めましたが、ど素人では手に入らない雑誌で、小保方さんの指導をしていて共著者のカレン博士がそれを自分のブログに公表してくれたのでやっと読めるようになった。なんだかひどい英文だと思いましたし、共著者に二人も米国人が居るのに、英文添削もしてくれないのかと思いましたが、彼らは英文には興味が無くて、報告されている事実にしか関心がないんですね。
三胚様分化したという事実だけです。これがESだったら当たり前の現象です。小保方さんはヴァカンティ研で3人の仲間と小島博士とカレン・ウェスターマン博士の指導の下に、分化確認をした。これはずっと後に小島さんが一部神経細胞分化は確認しています。
理研は再現実験時に三胚様分化確認はしたと思いますよね。常識的に小保方さんを疑うのだったらまずこの実験でしょう。この実験が嘘だったらキメラ確認なんて不要なんです。小保方さんを犯人だと決定できるではありませんか。それですべては終わる。ところが彼らはキメラ実験を行った。つまり、三胚様分化はしたのでしょうよ。三胚葉分化確認実験は2週間から6週間でできる。お金も大してかかりません。
三胚様分化確認実験を行って分化しなかったのにキメラ実験をすることはありません。問題は三胚葉確認実験もしないのにいきなりキメラ実験したのかということです。あり得ないと思いますがね。
小保方さんは2011年11月のキメラが出来る以前にメチル化確認実験を行っている。小保方さんの不正は二種類あって、キメラができているのだからこうであるべきだという実験事実の歪曲、もう一つがGFPが光っているのだからこうであるべきだという実験事実の歪曲で、メチル化実験はまだキメラは出来てない時点だということです。しかもこの時に若山さんの圧力があったことを桂報告は指摘している。
ここはOct4-GFPが光っているからメチル化解除されているはずだという強い思いがあって、それは若山さん自身にもあったんです。捏造しろなんて言うはずもないが、メチル化解除されているはずだぞということは言ってるはずです。このGFP蛍光が漏れ出しだなんて発想は誰にもありません。この時に、小保方さんがもちゃもちゃせずに、メチル化解除されてませんとダイレクトに報告したら、後に丹羽さんが確認した現象の発見に方向転換していたでしょう。小保方さんは自分の細胞がどんなに確率低くしか三胚葉分化しないかということを若山さん以上によく知っていたはずです。
ともあれ、ここで若山さんはこの蛍光細胞が何物かであることを強く信じた。そしてそれをntES化した。小保方さんがメチル化実験で不正をしなかったら、つまり、実験事実に忠実であったなら、若山さんはこの細胞をntES化してみようとは思わなかったかもしれない。それよりも、メチル化したままなのにどうしてOct4-GFPがこんなに光るのかという疑問に忠実な実験をしたでしょう。
舞い上がってるという感じがしますね。
7.ネイチャー誌のリジェクトを受けてのTCR再構成確認実験2012年8月になると若山さんは理研の知財と、自分に51%、小保方さんに39%、ヴァカンティと小島に5%ずつの、幹細胞株化の特許申請手続きを始めたと手記にある。GRASヘの細胞提出はこの特許申請のためのデータづくりです。後のレター論文のためではありません。笹井さんが参加するなんて話は8月には爪の先ほども無い。
この話は小保方さんが手記の中で勝手に言ってるだけですが、理研側からもヴァカンティ側からも否定されていない。ヴァカンティは小保方さんに直接「僕を騙したのかい」と問い、彼女は「5年間も捏造のために没頭しない」と答えていて、ヴァカンティは今も特許申請維持している。手記は米語に翻訳されていてヴァカンティも読むし、小島さんは日本人なんだから当然読んで、嘘があったらヴァカンティに言いますよ。それどころか、その後に三胚葉分化確認実験をしてネスチンを発見したのは小島さんで、小保方さんの話に嘘が無いことは確認していて、ヴァカンティも彼女を信用している。
尤もヴァカンティももうキメラが若山さんの工作だというとは知ってしまっていて、特許のクレームは最初申請しなかったティシュー論文の成果に狭められてしまっている。でも、ヴァカンティにとってはこの特許の戦いしか自分の無実を証明する手段が無いんですね。米国だったら警察が入って調査して裁判で決着します。日本では役所の中の自主調査と再発防止という法の立て付けなので、ヴァカンティは日本の曖昧な決着の結論によって、米国での常識的処分に自主的に応じている。つまり、長期休職している。とても困っているわけです。日本で裁判してくれたらどんなにうれしいでしょうね。しかたないから間接的に特許申請で自分の身の潔白を示すしかない。ただ特許は認められたらティシュー論文の真実は証明されますが、認められなかった場合は、論文の真偽とは別に新規性の問題がありますから、身の潔白に関しては灰色のままに残ってしまう。その時はヴァカンティ氏が若山さんを米国法で訴訟する可能性は残るでしょうね。
取り敢えず小保方日記では彼女はどこかの研究室に拾われて今静かにしていることになっていますが、特許の方が決着したらまだ事件は蒸し返される可能性を秘めている。国の機関であるような仄めかしですが、研究そのものがどうなっているでしょうね。三胚葉分化したという事実は重いものですが、この細胞が新規の多能性細胞であるかどうかは別問題です。
日記は2016/10/10で終わった。3年経過しています。ヴァカンティの申請はまだ続いている。
他方、私の仮説である小保方細胞核使用ntES実験はどうなったのか。胎盤蛍光はヴァカンティと小保方さんの細胞とは無関係で、単に若山さんのntESキメラの胎盤蛍光であった可能性が高いのでした。ここに笹井さんがさまざまな検証を残してくれた。これがレター論文なんですよね。でも、今はまだその検討まではいかない。まだ笹井さんは参加していない。小保方さんの論文が最初のネイチャー提出でリジェクトされて、西川さんがTCR再構成の確認をしてみろとアドヴァイズしたところです。
この時、小保方さんはまず蛍光してない細胞と蛍光している細胞でTCR再構成確認した。順番として当然です。どちらも同じCD45陽性細胞だ。ではSTAP幹細胞はどうか。この実験を若山さんは小保方さんにやらせなかった。
8.幹細胞に関して小保方さんに直接やらせなかったTCR再構成確認実験はネイチャーがリジェクトして、次のセル誌から記載されて、このことが書かれている。どういう書かれ方かは論文が無いので分からないが、最後のサイエンスの査読文は公表されていて、ここに、キメラのTCR結果に関してコメントされている。疑われている。
>>
The DNA analysis of the chimeric mice is the only piece of data that does not fit with the contamination theory. But the DNA fragments in the chimeras don’t look the same as those in the lymphocytes. This assay is not properly explained. If it is just an agarose gel then the small bands could be anything. Moreover this figure has been reconstructed. It is normal practice to insert thin white lines between lanes taken from different gels (lanes 3 and 6 are spliced in). Also I find the leading edge of the GL band suspiciously sharp in #2-#5.ここでつけられていた図が何かは分かりません。後のネイチャー再投稿時には幹細胞とキメラの結果は丹羽さんがはずさせていますからありません。ここで白線を入れる話をしているのがアーティクルの図表とは限らないばかりか違う構成になっている図でしょう。
我々の仮説では若山さんは小保方さんの論文は通ってもらっては困るんです。キメラは別の実験のものだ。高名な雑誌にアクセプトされて知れ渡ると自分のやったことが捏造になってしまう。だから落ちてもらわないといけない。でも論文は書かせないとヴァカンティが小保方さんを手放さない。小保方さんを山梨大に連れてきて、ちゃんと本当のことを言ってから幹細胞化実験を共同研究したいんです。人には自分が何をやっているのかは教えたくない。
彼女が山梨に来ると言ってくれたら、初めてヴァカンティにはESのコンタミ事故があったようだと説明したらいい。あきらめてくれるだろう。
彼女が来ると言わなかったらどうなるのか。そのときもどうも後で調べたらESコンタミだったみたいだから、キメラができたことは間違いだったとして自分たちで研究してくれたらいいよといえばいい。
それがよりによって理研で採用してしまうなんて。オホホポエムは身勝手な本音を語っている。
>>
「世に倦む日々の人、竹市のこと信用してたのね。御愁傷様」
「竹市は本件をここまで深刻にした張本人です」
9.幹細胞とキメラTCR結果が若山さんの予期した結果で無かった可能性でも、8月までの時点ではすべてが流動的ですね
①若山さんはまだ自分の研究すべてが終わっていない。
②TCR再構成確認結果はntESとしても予期しない結果ではないのか。
③小保方さんは8/21のサイエンス通知をもって3誌ともにリジェクトされた。でもまだ山梨に来ると言ってない。
④彼女が8月にGRAS提出した試料は若山さんの幹細胞化実験に絡むもので、自分の研究分ではない。
⑤手記では10月頃にラボ仲間も早く返事しないと言っている。若山さんはまだ誘い続けている。何しろ半年後には理研研究室は引き払って山梨で活動する。助手も早く決めないといけないのに彼女はまだ返事してない。
⑥11月10日頃に彼女はヴァカンティの許に帰った。
彼は4月初頭に胎盤が光ったのに小保方さんに免染を頼んだまま結果も聞かずにTS培地誘導実験を進めた。そしてCTSを作って、キメラ実験までしている。小保方さんにテラトーマ実験もさせている。
小保方さんは手記ではSTAP細胞由来キメラ胎盤をわたされているのだが、ここで「スフェアからのキメラマウスの胎盤だというものをいくつか渡され」という書き方をしている。酸浴細胞は長く培養維持できないので直前に作らされていたらそれと分かるが、ずっと以前のものだと凍結でもしておかないといけない。通常は帝王切開したての生ものである。小保方さんは事前に渡された赤ちゃんマウスの記憶が無かったのではないか。このことはなぜ2011/11/28のものとされるキメラ写真を免染時のキメラ写真としたのかと関係しているかもしれない。報告書はどうしてこれを調査しないのかな。両方に聞けばわかることだ。若山さんにでは渡した胎盤は何時のだと問えばいい。その時のキメラ写真は撮ったのか、渡したマウスの背景はと。どうして確認しないのか。どうして桃子に時期を言わなかったのかの理由とも連動している。
我々は小保方核使用ntESだと考えている。しかし、ここで仮に太田ESだったとして、胎盤が光るのは全部嘘だと。若山さんが騙されたのだと。しかし、騙された結果、太田ESをTS培地誘導したらCTSができて、そこからキメラとテラトーマもできた。胎児と胎盤のつなげたままのものもある。木星リストにある。
これらはすべて太田ESキメラだったのだと。胎盤は光ってないのだと。桂報告の推理はそういうことになる。
太田ESはTS培地で維持できるんだな。以下のプロトコルは若山さんが教えたもののはずだ。
>>
FI stem cell conversion culture
1. STAP cell clusters were transferred to Fgf4-containing trophoblast stem-cell medium (Tanaka et al, Science, 1998) on MEF feeder cells in 96-well plates (Obokata, Nature, 2014b).
IMPORTANT
(i) TS medium consists of RPMI 1640 with 20% FBS, 1 mM Sodium Pyruvate, 100 µM 2-mercaptoethanol, 2 mM L-glutamine, 25 ng/ml of recombinant FGF4, and 1 µg/ml of heparin.
(ii) Different lots of FBS may results in significant differences in the behavior of cultured cells.
2. In most cases (40 of 50 experiments), colonies grew in 10–50% of wells in 96-well plates. In a minority of cases (10 of 50 experiments), no colony growth was observed and/or only fibroblast-like cells appeared.
IMPORTANT
(i) The cells in proliferative colonies also appear similar to fibroblasts, but gradually change morphology, coming to resemble epithelial cells.
3. The cells were subjected to the first passage during days 7–10 using a conventional trypsin method. Subsequent passages were performed at a split ratio of 1:4 every third day before they reached subconfluency.
IMPORTANT
The cells must not be dissociated completely. Partial dissociation is optimal to maintain viability and self-renewal, as seen in the case of embryo-derived trophoblast stem cells.丹羽さんはこの培地はTSに適合的な培地だと書いている。むろん、小保方さんの細胞では樹立できない。桂報告書は太田FES1からなら樹立できると言ってることになるので、どうして樹立確認しなかったのか不思議ですね。簡単だよね。FES1は太田さんのところにあるし、培地はプロトコルに書かれている。残されたFES1ではだめだよね。FLSに中身が入れ替えられている。
FES1、2は受精卵ESだということになってますからね。胎盤は光らないよね。木星リスト-30度フリーザーの14~17番にFI-SCキメラの胎盤羊膜胎児の切片スライドがあるぜ。どうして調べないの?
このFI-SCに関して笹井さんが後にとんでもない検証実験を行ったことになってしまった。JAK阻害剤とBFPを使っていろいろと調べた。ただ、それはこの1年後の話で、この8月のAC129の実験の裏に有りそうな実験の真の目的が何か。2012/8/12に培養開始されている。FI-SCの実験は6,7月の実験ですね。その後にAC129の実験を行った。
サイエンスリジェクト通知は2012/8/21です。
(L氏コメントチェック)わたしは常時チェックしているんですよ。
5627. L
2019年11月24日 03:34
割り込みすいません。大筋同意しますが、多少言い過ぎな気もするので。
>Oct4発現までは進んだ研究者もいましたが、調べたところ自家蛍光であると判明し、結果的にそこに参加していたすべての実験で再現できずでした。検証実験でも同様の結果、見解でした。
GFPレポーターでOct4プロモーター活性を検出できたのは、検証実験の筆頭著者だけです。他は、丹羽先生やデイリーを含め、いずれも自家蛍光との結論でした。 Oct4-GFPの再現は世界各国で不可能だったわけですが、検証実験は厳密に管理されており 、FACSを担当したのは筆頭著者ではないので、再現不能でも不正とは言えない事はご理解頂けると思います。世界のトップラボで再現できないからといって、そのような現象(酸浴でOct4-GFPが光る)が存在しないとは断定できない一つの事例であり、再現性だけで評価する事の危険性を感じます。
再現しなかった理由として、筆頭著者が何らかの技術を持ち合わせていた可能性がありますが、ご本人がそれを自覚していないか、きちんと詰めきれていなかったため、ブラックボックスになってますね。ちょっと不幸な事です。
もう一つの可能性として挙げられるのは、 使われたマウスの問題です。デイリーのラボのマウスは、Oct4プロモーターに改変が入っており、ナイーブ型ES細胞レベルまで初期化されないと光らないようになっています。プライム型ES細胞レベルの不十分な初期化でも光る理研のマウスとは異なっており、再現性を論じる上では問題と思われます。学さんの言い分にも一理あると思います。
5628. L
2019年11月24日 03:38
>検証実験や追試の結果から、彼女にはOct4を発現する細胞隗までは作れていたのでしょう。しかしその先の多能性確認で思ったような結果が出なかったので捏造、改ざんしてデータをそろえたと思われます。
キメラのデータがあったので、 「Oct4を発現する細胞隗までは作れていた上に、キメラまで作成されたため、多能性があるとの前提に立ったサポートデータを集める必要があり、その要請に応えるために不正でデータをそろえた。」くらいまでしか言えません。それでも不正は不正ですけどね。ES混入ルートが不明という制約下では、これ以上言えないですよね。
>小保方さんが不正をしないで誠実に真正のデータを残し、自家蛍光であると見極められていたら
適切なネガコンを設定して、自家蛍光をきちんと識別できる実験系を組んでいれば、不正の有無に関わらず、きちんと見極める事ができたと思います。この責任(実験系の不備)を、筆頭著者一人に負わすのか、チームで負うのか、という事です。
筆頭著者が不正をしないで誠実に真正なデータを残した場合でも、自家蛍光の誤認がそのままだと、Oct4-GFP偽陽性に基づいてキメラの実験に進んだと思われます。そこでES混入がおきれば、チーム全体で「行ける」という思い込みで突き進んでしまう可能性が高いと思います。Oct4-GFPのところでプロジェクトを止めるのに必要だったのは、真正なデータではなく、それが自家蛍光である事を判定できる実験系だったという事です。①デイリーのラボのマウスは、Oct4プロモーターに改変が入っており、ナイーブ型ES細胞レベルまで初期化されないと光らないようになっています。丹羽さんの使ったGOFも改変型だったのかな。
どういう仕組みでESレベルまでプロモーターが働いていることを認知する仕組みになってるのか解説を乞う。
丹羽さんのGFP漏れ出し現象に関して解説願いたい。自家蛍光との関係の説明。
因みに、小保方さんの細胞は丹羽実験でもOct4タンパク発現は確認されている。また、その細胞のトレースもローザGFPで確認されている。
********
丹羽さんの写真をつけようとして書き込み禁止になってしまって元のブログを破棄しました。Ooboeさんのコメントをここ本文に移しておきます。
コメント
分身さん達、お久しぶりです
Lさんとのお願いについての思い残念だけど了解です。
もう少ししたら、パートナーは
差し支えない範囲で
楠本さんに検察申告の件資料を順次UPしてもらう準備をしているところです。
2019/11/14(木) 21:22:28 | URL | #- [ 編集 ]
- 2019/11/25(月) 09:25:13|
- AC129
-
-
| コメント:0
ガンバレ西岡さんブログで楠本さんがデータを貼り付けておられる。こちらにも保存しておきましょう。笹井さん記者会見時資料ですね。
簡単に解説しておきましょう。
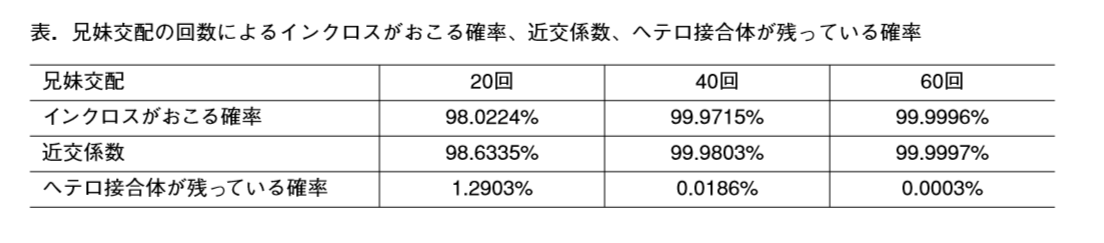
「STAP現象を前提にしないと容易に説明できない部分」とある。
①ライブセルイメージングでGFPの蛍光しない状態から蛍光する状態に移行したことが観測されたこと。
②細胞の大きさが違うこと。
③キメラができたこと。
以下がその詳細説明です。
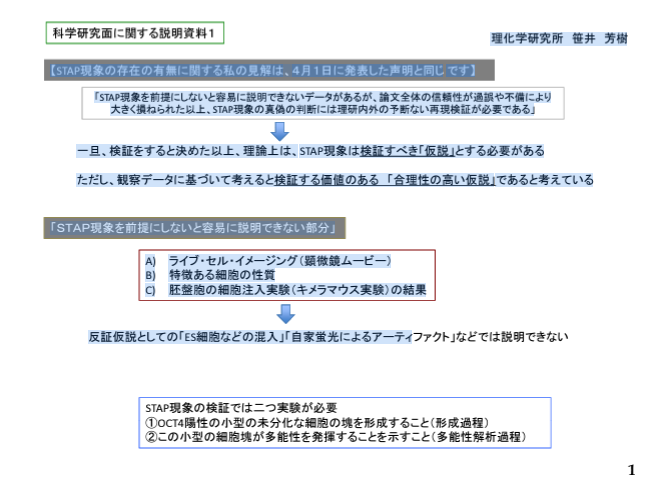
①に関して言えば、ESコンタミでないのは自明で、最初光ってない。撮影は全自動で途中で止められない。自家蛍光でないのは多画面のフィルター確認は無論のこと、ここではFACSでも確認されていることをあげているが、私は自家蛍光が6時間しか持続しないことと、この撮影のインターヴァル計算からも証明している。
これに関して丹羽さんの検証実験でOct4-GFPの「漏れ出し蛍光」があるという新事実の指摘があったんです。誰も知らなかった科学的事実です。Oct4-GFPの設計者は今とても関心を持って確認しているのではないですか。
②に関しては、笹井さんはES研究の大家でESの大きさは知っているんです。ここには小保方さんがArticle Figure 1-gに添付した画像を笹井さんはそのまま比較している。でも小保方さんは自分でも言っているように実験事実を分かりやすく提示するのが画像の役目と心得ている。ここでは形の違いを示すためにCD45、STAP、ESのほぼ全体の形が見える大きさに揃えて比較している。彼女のとても悪い癖ですね。出来るだけ事実に近く表示すべきです。左下にスケールバーがあって、写真の大きさを変えて三枚一組にしているのが分かる。意図は形の違いです。大きさの違いを示すのなら左下のスケールバーの長さをそろえなければならない。3枚組写真を示している意図が違うんですね。こういうのはとても誤解を受けやすいでしょうね。なぜ素直にそのまま図を作らないのか、自分の説明の意図に従って分かりやすくしたいのです。捏造意図ではないことは無論で、スケールを一致させると大きさの違いはもっと際立ちます。
③胎盤の光ったキメラに対してマスコミの騒いだような捏造ならESとTSと混ぜることになるが、細胞は接着しないと言ってる。このことは丹羽さんが過去に自分でやってみたことがあったんですね。丹羽さんはES細胞の胎盤研究をしていて、2005年にいろいろと遺伝子発現の違うES細胞の選別を行って胎盤を蛍光させるのに成功している。むろん、小保方さんにはそんな技術はありません。丹羽さんはいろいろと試行錯誤している時にESとTSの共培養もしてみているんですね。培養液が違う細胞なので難しくて、三回目くらいの継代で消滅してしまうと説明していましたね。
そもそも、TSは2012/5/25の培養開始でしたよね。小保方さんにSTAP細胞の胎盤と言って渡したとされる時期は3月の終わりから4月の始めです。小保方さんはTSを持っていない。
桂報告書では胎盤は光ってないという結論になってます。でも、胎盤蛍光写真は二つほどありますからね。一つには免染画像がつけられている。報告書はそもそも光ったことにしたかったという動機の面を指摘している。推測です。そして画像は若山さんは嘘だと言ってるが、露出の工作は無いと言っていて、しかし、免染写真については、そもそも光っておかしくないものを光ったと言ってるだけではないかとこれも推測している。
加えて、桃子本の中で若山さんはESも技術の進歩で光るようになっていたかもしれないと答えていて、この時ESのことを言ってるのに、自分の研究室ではキメラ達成率の向上を目指していてと、おいおい、それはntESじゃないかという疑義を掛けられても仕方が無いような口の滑らせ方をしている。
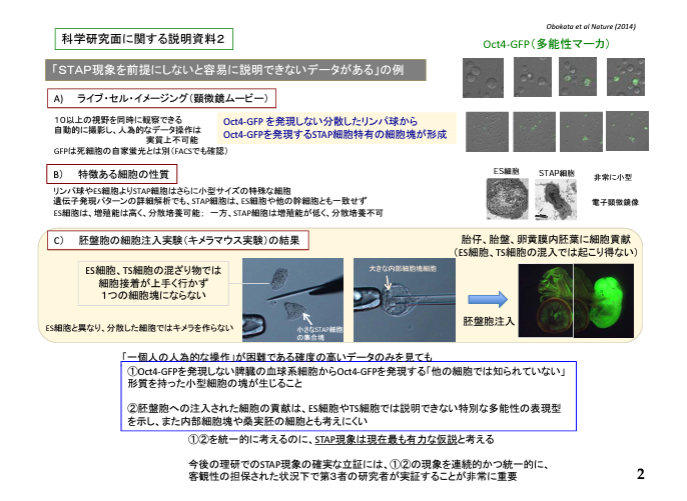
笹井さんは若山さんが小保方さんの細胞の核を使ってntESを作っていたんだなんて思いもよらないことなんですね。彼は最後まで論文通りのSTAP現象を信じていた。若山さんが何かしたとは爪の先ほども疑ってません。
ただ、笹井さんはSTAP幹細胞がどうして小さかったはずのSTAP細胞を培養している過程でES並みに大きくなったのかということに気づきませんでしたね。
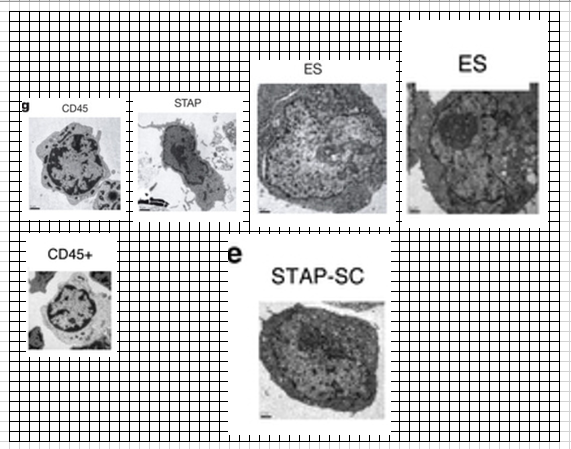
酸浴STAP細胞の核を抜いてクローン胚に入れntESとして培養したから大きくなったんですよね。
彼はこの後FLSのジャームライン確認実験に入り、恐らくFLS(実態は小保方核使用ntES)のキメラの胎盤蛍光を発見し、そのTS培地誘導を考えた。ただし、小保方さんはまだ自分の細胞からのキメラ作成だと信じていてその論文を書いているので、その実験を通して別の実験を行っている。
AC129-10に戻りましょう。
- 2019/11/23(土) 09:59:43|
- AC129
-
-
| コメント:0
Ooboeさんから連絡が来たので、ちょっと中断してお返事しましょう。なにしろOoboeさんとは以前村松のフルートの話で盛り上がって以来の親友なので、こんな中断は意にも介しません。僕の分身たちも大喜びです。
小野小町
Ooboe さん、お久しぶりね。お元気ぃ。金髪さんもジムさんのところには見えるのにここには見えないから寂しいわね。いつもハワイアンコナを持ってきてくれてたのに、キラウェアが噴火してからきっと入手困難なのね。うふふ。
在原業平
今度又奈良に行きます。前回見れなかったところと、足を延ばして伊勢神宮にも行くつもりです。このブログで又写真アップします。
小野小町
取り敢えずあなたのコメントは以下に貼りつけるわ。
>>
居士さんのお陰でパートナーは質問の資料の整理ができました、ありがとうございました。
さて、パートナーと理研広報や情報公開室を介した調査委員会事務局とのやり取り(CD録音)により、複数の理研組織において、サンプル出所についての認識を、それぞれ組織が同一の報告を共有していたことが明確になってますが、これらの報告資料ではサンプル出所はFES1とntESG1は 山梨、若山氏から、FES2とntESG2は 別の処から、に対しBCA論文ではFES1と2、ntESG1とG2は太田浩から と矛盾。
この矛盾資料の確認質問書を京大、太田氏と桂調査委員に送付しました。併せて、居士さんに教えていただいたオス、メスの矛盾指摘のため、該当ページ論文と、検察への申告書も同封しました。
2019/11/12(火) 08:46:07
それから、検察には、以前の手続きでは提出できなかった、オス、メス矛盾資料をお陰さまで提出します。また今回の委員や太田氏あての質問書送付についても報告するとのことです。
2019/11/12(火) 09:18:11
居士さんのこと、すでに閲覧されているかも知れませんが、Lさんが、このAC129考察についてコメントされてます。私達は、居士さんのスピン評価のようには、彼のことを受けとめていません。
彼は、居士さんと同じく科学的にニュートラルなスタンスの方と思います。
ただ時系列的把握や小保方資料を文献的に認識する作業が海外ての研究制約のため時間的に不十分のようです。断片的なので誤解されやすいかもです。
トータルな把握がなされ(ここ切れてますので後で追加修正してくださいね。)
2019/11/13(水) 09:54:34
それから、お願いがあります、居士さんなら素人の代表として、研究者のLさんとの、純粋なスタンスでの科学議論が成立すると存知ます。結論ありきの雑談コーナでお願いします。
2019/11/13(水) 10:27:55
私は、雑談コーナに一度コメントしましたが、以後入れない判断をされたようです。
2019/11/13(水) 10:32:53鉄
相変わらず名前を入れないんだな。
ふふふ
鉄、Ooboeさんはなあ、スマートフォンだから入力が大変なんだよ。いちいちチイセエことに目くじら立ててんじゃねえや。
鉄
親分、俺たちは以前[「結論ありき」の非専門家ブログが社会を混乱させている:世界三大不正STAP事件の正しい理解を社会に広める会]という人間の屑たちの集まっているところに一度投稿しませんでしたかねえ。
ふふふ
そうだなあ、俺たちは一度[「結論ありき」の非専門家ブログが社会を混乱させている:世界三大不正STAP事件の正しい理解を社会に広める会]という人間の屑たちの集まっているところに一度投稿したよなあ。アク禁くらったっけなあ?
閲覧者
いやいや、我々はかつて[「結論ありき」の非専門家ブログが社会を混乱させている:世界三大不正STAP事件の正しい理解を社会に広める会]という人間の屑たちの集まっているところに一度投稿したけど、アク禁食らうほど長くはいなかったよ。あそこは臭くて長くは居れないところだよ。
小野小町
Ooboe さんったら、ちゃんと相手の家の門の表札見てる?[STAP細胞(実際には小保方氏の虚構)]と書かれているのを見てないの?
>>
「世界三大不正」の1つとも呼ばれる、STAP細胞ねつ造事件の正しい理解を社会に広めることが急務だと思い作成致しました。 現在この件について、科学的見地から正しい説明をしようとする研究者および良心的な方々に対し、STAP細胞(実際には小保方氏の虚構)があるものと思い込んだ人々からの脅迫行為なども行われています。その結果、良心的な研究者がほとんどSTAP細胞について発言できなくなりました。この異常事態を改善し、STAP細胞についての正しい知識を社会に広めるため、このブログを立ち上げました。鉄
ひひひひひ。
STAP細胞は若山氏の虚構だよな。
一言居士
Ooboe さん、あそこは感想さんの立ち上げているブログでしょ。あれは若山研に居たテクニシャンじゃなかったですか。李氏とも知り合いで、ノフラー氏ともハァーイかなんか言っちゃって無かったですか。大阪の万博館かどこかにいらっしたのかな。
鉄
あんなところに書き込んでるやつらなんて全部スピン屋だよ。いつも同じメンバーじゃないか。金もらってやってる。仕事なんだから邪魔しちゃいけないぜ。人は生きて行かないといけない。
ふふふ
まあ、そういうことで御期待には添えませんが、我々は常時チェックはしてるんです。何れことと次第によっては摘発されるかもしれない連中ですからな。我々のしたらば掲示板をああやって破壊した奴も連中の仲間です。我々は復讐を誓ったらとことんやる人間なんですよ。
鉄
親分、モンテクリスト伯みたい。ひひひ。釣りしながら復讐心を煮えたぎらせてるんだな。
ふふふ
あ、雨だと言ってたのに天気になったな。ちと、釣りに行ってくる。
小野小町
あら、ふふふさん、お帰りなさい。マブの入れ食いだったんですって? あはははは。
鉄
親分。今回の釣り紀行の写真はいけねえや。
ふふふ
なんで?
開高健
あの場所、分かるぜ。
アイザック・ウォルトン
君、STUDY TO BE QUIET.って僕が教えて置いたはずなんだけどなあ。
鉄
親分。復讐心でフツフツと心が煮えたぎってたんですね。あらぬこと考えてっから。ひひひひひ。
ふふふ
そうでもないよ。あんまり釣れないから早々と切り上げて周辺の草刈りしてた。
山村聰
釣れない時って帰るに帰れなくて要らん事するよなあ。
閲覧者
Ooboe さんのパートナー氏は着々と計画を前に進めておられるな。
一言居士
僕はCDを聞いてないから何とも言えないんだけど、BCA報告がFES1、2、ntESG1、G2の4つのサンプルを太田浩氏からと言ってるのは、ただ、これら4つの細胞は太田さんが作ってすべて京都大学に持ち出したと言っている以上、経由地点はともかくとしてすべて太田氏だと論理的に推測しているだけではないかな。出典はExtended Data Table 1だね。
>>
Obtained from H. Ohta. Full names: FES1: 129B6GFP1 FES male; FES2: 129B6GFP2 FES male; ntESG1: 129B6F1G1; ntESG2: 129B6F1G2.
閲覧者
桂報告書は少しニュアンスが違うね。
>>
STAP 幹細胞 FLS が、Acrosin プロモーター下に GFP を発現する Acr-GFP と、CAG プロモーター下に GFP を発現する CAG-GFP の共挿入を含むことが判明した後に、 過去に CDB ゲノム・リプログラミング研究チーム(以下「CDB 若山研」という)で作製さ れた Acr-GFP/CAG-GFP 共挿入 ES 細胞を取り寄せて解析したのが、下段の 4 種類の細胞である。小野小町
誰が何をして、どういう経路で、最終的に理研に来たのかが書かれてない。
一言居士
我々は戦略的にパートナー氏は情報を全開示していないと思ってるけどね。お任せしておきましょう。
小野小町
それで何? Lさんの話がどうなのよ。一応貼り付けましょうかね。
>>
5557. L 2019年11月12日 10:29
一言さんが、長い記事をあげてます。どなたか反論ありますか?
BCAで記載されているFES1とFLSのギャップ(獲得変異の不一致率30%)が、感想さんの解析では見られない(9割以上の獲得変異がFES1とFLSの間で一致)事に絡めて、一言さんの主張について考えてみたい気もするのですが、長い記事を細かく読む暇がありません。
ちなみに、一言さんが「残された疑問」としている3点のうち、1番と3番は解決済と思いますよ。CAG-GFP内にプライマーを設定したinverse PCRで挿入部位を同定し、捕まったアクロシンプロモータを内在性と誤解(実際にはコンストラクト内にある)して15番としたのでしょう。18番はもっとシンプルで、実験4でPCRがかかって、実験3でかからなければ、このlocusにはCAG-GFPなしでいいでしょう。
3番目の問題は、キメラ子の親で配偶子ができる過程で、減数分裂組み換えが起きて、Acr/CAG-GFPと欠失が分離したという事でしょう。
どのような意図でローザと説明したのかは不明なので、この問題は残りますね。 在原業平
Ooboe さんは賑やかしの好きな人だからねえ。でもこれってLという人の感想でしょ。人の感想には興味ないねえ。僕は忙しいんだ。
鉄
「長い記事を細かく読む暇が」無いんだろ。仕事せえや。若いうちからさぼり癖をつけるな。死ぬほど働いたこともねえんだろ。クソが。
ふふふ
鉄、お前は口汚くていけないよ。留学して何をしているのかは知らないが、誰の金でイギリスに行ってるのかな。親が泣いてるのか、国が泣いているのかは分からないが、若いうちの時間は濃密だからな。無為に流れたか身になったかでは後のレバレッジが違うよな。
小野小町
ふふふさん月末には奈良に行くんでしょ。早くホテルの予約しないと。コース決めなさいな。
ふふふ
法事の写真も送んないといけないんだったな。何しろ年取ると一日が短いのにやりたいことが多すぎて、そしてすぐ疲れる。どうしたらいいんだ。
鉄
親分。死ぬほど楽は無かりけり。もう少しの辛抱ですぜ。
ふふふ
鉄、お前間違ってるから。寝るほど楽は無かりけりだ。そうだ。家に帰って寝よう。
小野小町
もう寝ちゃうの。まだ宵の口だというのに。
アイザック・ウォルトン
釣れなかった日の心は重い。
井伏鱒二
さぞかし瞼も重たかろう。
- 2019/11/14(木) 09:41:48|
- AC129
-
-
| コメント:1
(本題に戻ります)問題はSNPs解析に戻ります。再掲しましょう。
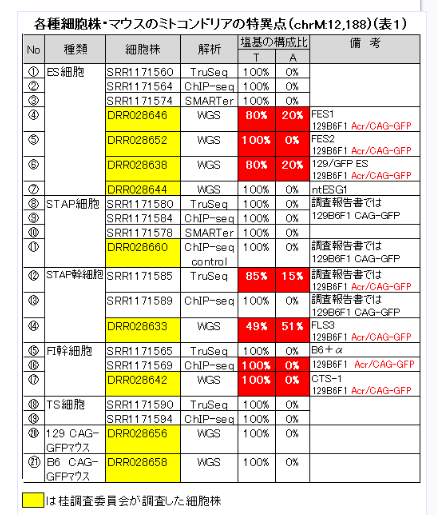
一番下のたまたま分析を受けた129 cag-GFP mouseの15番染色体のSNPsパターンは最上段3つの細胞とは違うというところを確認してください。これは若山さんの129マウスにマウスコンタミがあるからでしたね。解析を受けた129と上3つを作った時の129が少し違うのです。でも上三つは同じマウスからできている。そしてAC129はどうせ若山さんが「僕のマウス」ES-6を詰め替えたものです。しかし、STAP Lysateは違いますよね。これは小保方さんがChIP検査のためにかき集めた細胞だと証言していると伊藤さんが記者会見で語っています。人から取り戻している細胞ですね。このときに誰かが129B6F1 ES6を渡しましたね。AC129の実験時ではありません。小保方さんはF1のSTAP細胞として提出しているのです。
このことは何を意味するかというと、既に2012年の8月には小保方さんは嵌められているということです。そして原因は恐らく特許での対立です。若山さんはこの対立での経緯でこのまま放置できなくなった。万が一の時には小保方さんの捏造で逃げる準備が行われている。と考えると何となく辻褄はあう。でも、これはSNPs解析結果ですね。思い出してください。Nanogの発現解析はSTAP細胞とES細胞は違っていましたよね。伊藤さん。本当にSTAP Lysateが残っていましたか?
>>
ただ、唯一やったのが、あ、GRASの方に残っていたChIP seq のinput 、しかもSTAP細胞のインプットはDNAとして残っていたので、それが10ページバワーポイントの、えーと、ところに書いてある、3番のアイテマイズして書いてあるところですけれども、これに関しては、えーと、30倍になるだけ、他のゲノムと同量となるまで読んでいます。そして、読んだ結果、この、同一であるということを、ま、認定したということで、それ以外に、それ以外に関しては、ま、残っていませんのでやってない、やってません。(*ttps://www.youtube.com/watch?v=M9oJGioHvIQ 1:15:00から)
因みにここで伊藤さんはこのたくさん集めた細胞は別の実験だと言ってますね。つまり彼らはこれがAC129の実験時のSTAP細胞ではないことを聞いているんです。にもかかわらず、「公開データ再解析の結果に よれば、論文に記載された実験の中では Letter Fig.4 に使われた可能性が高く」と書いているのです。嘘ですね。彼らはここで小保方さんが若山さんの「僕のマウス」ESを使ったと主張しているのです。そのくせNanogの件は無視している。それならこの時のESは何なのか。
GRASに残されている記録を全部発表してもらいたい。若山さんのこの頃の実験ノートを出してもらいたい。どうして小保方さんばかりに訳の分からない因縁をつけているんでしょう。小保方さんは調査チームを信用して実験ノートを提出しているのに、彼らは全コピーをNHKに違法に流出させて犯人の検挙もしてないだろうが。告発すらしていない。犯罪を知ったら警察に届けるという公務員義務違反をしていて恬として恥じない。日本の知的レヴェルは明らかに下がってますね。こんなのが博士だと思うと情けないね。ははは。
さて、いよいよこの時の129/Svは何なのかという問題に入れるわけです。FLS3以下5つの細胞株の6番染色体が一番下の129 cag-GFP mouseとは違うことを確認してください。これも又別のコンタミマウスなんですね。でも他のB6のSNPsパターンは全部同じだということも確認してください。これって市販の129/Svマウスではありませんよね。アルイミオウジ氏も疑義を呈している。でももし129 cag-GFP mouseだったら18番染色体にヘテロでGFPが入っているはずです。
やっと最初の問いに戻りましたね。どうしてFES3のGFPは岡部マウス側のAcr-CAGのみが調べられて、母親側の18番にCAGが無いかどうかを確認されてないのか。もしそれがあったらFES1,2には無いはずのものですよね。仮にterで作ったという記憶が間違いでX1であったとして、しかもそれは市販の129/Svではなく、更に若山さんの129/Sv-CAGホモだったとして、更に、どうして7年も前のコンタミマウスのSNPsパターンの同じ親にぶち当たるのだ。
FES1,2ともに中身の細胞は若山さんによってFLS3に入れ替えられているということです。
さあ、大問題です。若山記者会見です。
*ttps://www.youtube.com/watch?v=hD3Wd9cpo78
FLS1-8は15番にヘテロで4コピーのGFPがあり18番には無い。「僕のマウス」シリーズには15番にはGFPは無く、18番にホモで1コピー入っている。誰でも知ってることですね。
ところで15番にGFPが入っているという第三者機関の発表は間違いとされた。遠藤さんが発見したことになっているが、それはともかくとしてではFLSの18番にはGFPが無いという結論はどうなっているのか。一方が間違っているのに他方は合っているというのも変な話しで、その場合は間違った原因を述べなければならず、又同時に18番を調べた手法の確認も必要です。でもそれはうやむやになっているままなんですね。SNPsパターンは129x1/Sv-CAGホモマウスの特徴がほとんど全染色体に分布してますよね。どうなってるんでしょうかね。
今、アルイミオウジ氏がいくら兄妹交配を重ねても決してホモ化しないSNPsがあるなどというトンデモ主張中ですが、又、テヘで終わるのでしょうかね。そんな現象があるのなら、そもそも報告書のSNPs解析は意味を持ちませんね。
有る特定の129マウスが近交系マウスになったというのは兄妹交配回数が20回を超えて、基本すべての対立遺伝子がホモに揃っているものです。129特異的SNPsというのはその全遺伝子の並びの中で野生型と比較してSNPとして違っている箇所が特定されているということです。近交系マウスの中では全部ホモになっているSNPsです。これは定義なので近交系マウスの中のヘテロなSNPsというのはあり得ないんです。まずヘテロなSNPsになっていたら近交系マウスではない。以前から説明せずに一人よがりする人ですから他所から見て何とも言えませんが、近交系マウスの中でヘテロなローカスというのはいくらでもある。全遺伝子がホモに揃っているというのは実験目的での実用の範囲で言うものです。実際にはヘテロなアレルは残っています。以下が参考になる。
*www.anim.med.kyoto-u.ac.jp/Kuramoto/contents/ExpAnimGenet_1_inbred.pdf
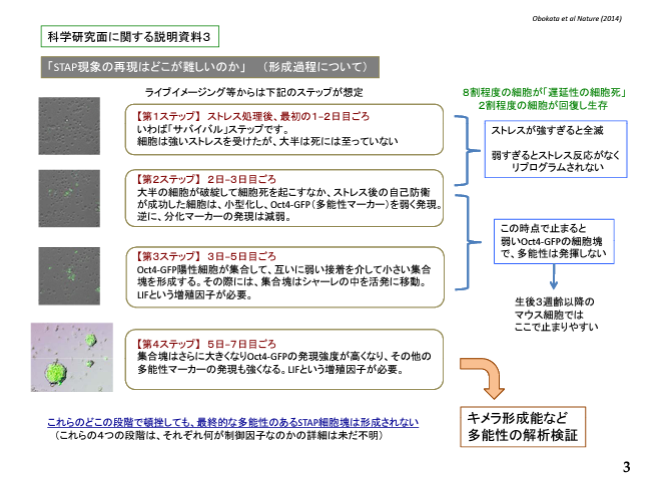
マウスの場合は胎児から成熟して妊娠できるようになるまで20日と50日の合計70日かかる。20回の交配というのは2.8年、40回の交配というのは7.7年、60回の交配は11.5年です。マウスの全塩基数は30億個程度です。60回やってヘテロ残存率が0.0003%になった時の数は9000個です。染色体数を20として一本に450か所ある。20回の交配後既に近交系マウスとして扱われるので、その場合では38,709,000か所残っているので染色体1本につき約2百万か所のヘテロなアレルがあることになる。
このヘテロなアレルは両親の違いがそのまま残っているだけのものです。これを更に兄妹交配して世代を重ねていくとヘテロだったものがどちらかに固定されていくんです。一旦固定されると何度交配してもホモなんですから子供には同じ核酸が分配される。これがインクロスと呼ばれる交配なんです。全部がインクロスになったら完全クローンになりますが、理論的にはこのやり方では無限に繰り返してもヘテロはわずかに残ってゼロに限りなく近づくが完全なゼロにはならない。
この問題と、ある近交系マウスに特異的なSNPsというのは無関係です。これは野生型と違う核酸になっている場所のことで、近交系マウスではほぼすべてのアレルがホモですから、ホモの中で野生型と違っている場所という意味です。ヘテロなSNPsなんて子供を造ったら分かれてしまうのでその近交系マウスに特異的SNPということにならないんです。定義の問題ですね。ホモで且つ野生型でない場所。そして近交系マウスの種類によって核酸の種類も場所も違っているものを特異的SNPsという。
まあ相変わらず一人で分かった気になってまともな説明が無いので端からは何とも言えませんが、この問題の解にはなってない筈です。
アルイミオウジ説の話になったので序に書いて置きますが、彼はntESG1,G2のGをgenerationと解して、最初のntESがG1で、それを使ってキメラマウスを作って更にntES化したのがG2と推測している。そして実際に若山さんはこのやり方で20世まで作っていて別の論文があるわけです。でも太田さんのG1ははっきりしていて、2008年論文で2005年論文で作ったF1のntESを解凍して使った。その解凍した細胞を129B6F1G1と書いているんです。すると解凍する前の状態のラベルはどう書いてあるのかということになる。株分けした時に以前の分に129B6F1G2と書いたとしたらこれはgenerationの意味ではありません。ただ、日経サイエンスに太田さんは自分は2種類の受精卵ESと4種類のntESを作ったと証言している。太田さんに聞けばわかることで、彼には説明義務がありますよね。でも、チューブのラベルと中身が違っていることの説明は彼一人の判断では出来そうもありませんよね。
(残された問題の整理)さて、問題の数も少なくなってきましたね。ただし、解の困難さは増してきました。ここらで残されている問題を整理しておきましょう。
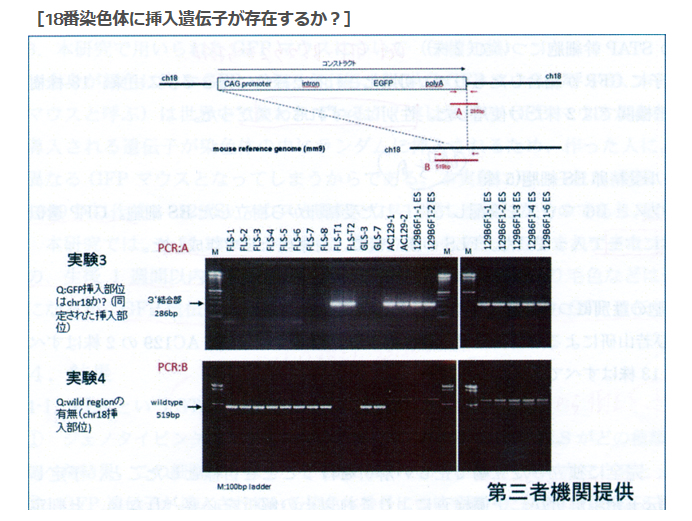
①FLSは「僕のマウス」を渡したと一方的に証拠の提出もなく若山さんが主張しているが、実際の検査で出て来たのはAcr-CAGのB6(岡部マウス)と市販のGFP無しの129X1/SvのF1だという報告書の調査結果となっている。しかし、129側の18番染色体に1コピーのCAG-GFPが入っているのではないかという疑義がでた。なぜなら、SNPs解析結果がこの129は「僕のマウス」の片親であることを示唆しているからだ。最初に放医研がFLSを検査した時、15番にヘテロに4コピーのCAGがあり、18番にはGFPが無かったと発表していたが、後に15番のGFPが否定されたが18番に関しては何も触れられなかった。又間違いの原因もちゃんと説明されていない。内在性アクロシン遺伝子を間違えたのならホモだと思うはずでヘテロに間違うというのは考えにくい。調査自体がとてもあやしい。すると18番に無かったというのは本当なのかという疑義が残るということです。理研のチームはGFPを自分でちゃんと確認したかということです。この説明も無い。
②小保方さんはなぜローザだという説明を受けたのか。彼女は徒手空拳で何の理由もなくローザと書く動機が無い。誰かがローザだといった筈である。
③なぜカルスキメラ子の8番に3番染色体欠失があるのにAcr-GFPがないのか。多分これが我々素人にとっては最大の難問になりそうですね。というのも他のことは誰かが何かをしたとか、し忘れたとかいう可能性を思いつけるが、この問題だけはど素人ではなぜこうなるのかの一つの可能性すら思いつけない。
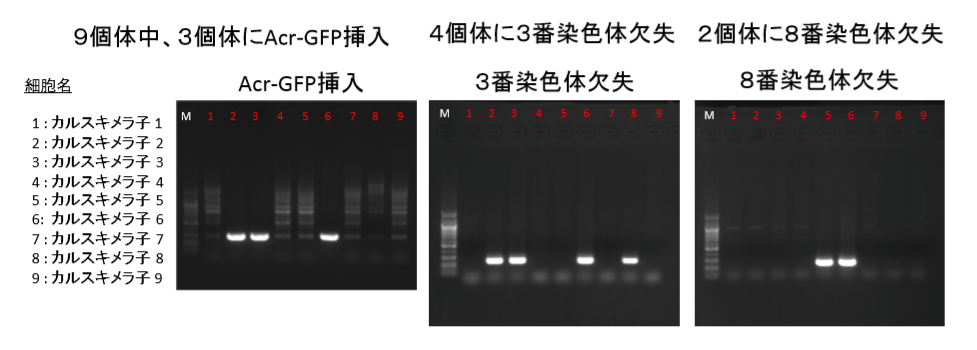
欠失とAcr-GFPはB6の同じ3番染色体上にある。一つの細胞に欠失が起きてそれが分裂して増える。最初は一つの細胞です。ここにAcr-CAGがあったらその後の体細胞分裂でGFPはずっと引き継がれる。欠失は同じなのにGFPが無いということはあり得ないんで、その場合は欠失の種類が違うということしか考えられない。逆にGFPありとGFP無しの細胞に同じ欠失が起こることも無いですね。まあ、せいぜい突然変異でGFPが消えたというなら分かりますけどね。そんな確率で起きることですかね。
この表ではB6に129がコンタミしている場所を指摘していますが、129にB6がコンタミしている場所を提示していませんね。もう一段深めて推定する発想力が無いんですね。そういう発想力があったらマウスコンタミの無いB6と129のデータをコントロールとして並べますよね。客観的たろうという姿勢が無いんです。まあ、科学者としての資質としては相当低い人々でしょうね。一般の会社に入っても役だたずでしょう。こういう奴って悪いことばっかり考えるんだよなあ。ときどき見かけるタイプ。流出させてはいけないデータを羽織ゴロツキとかNHKなんかに流す。
NHKに調査チームが病床の小保方さんの手元から奪い取るように持ち去った実験ノート3冊分の全コピーが流出するという犯罪があった。誰が流出させたのかをNHKが答える義務はない。又理研内での自己点検報告の草稿が作られたとき、それが毎日新聞の記者にリークされた。このリーク者を毎日新聞が公表する義務はない。しかし、二つの公務員としての犯罪が行われたことが明確であるにも関わらず理研がこの事件を警察にとどけることが無かった。これはこれ自体が犯罪ですね。
これは追及されるべき事柄です。日本は法治国家では無いのか。
内部告発も取材源秘匿の自由もちゃんと保証されている。でも違法行為は警察が取り締まるという義務も又課せられているんだとということを忘れてもらっては困る。理研は今からでもNHKに小保方さんの実験ノートを流出させた犯人を捕まえるよう依頼しないといけない法律になっている。無論、その際でも、NHKは警察に対して犯人が誰かを教えなければならないというような義務はありません。取材源の秘匿は法で保護されている。ただし、裁判所の令状があると別ですけどね。裁判所は裁判所で個々のケースで令状を出すか否かは判断します。このケースでは出ませんね。でも警察はそんなもの無くても犯人をすぐ特定しますよ。捜査権限がありますからね。実験ノートに関与した者なんて限られたわずかな人数です。こんなことをそのような人間が調査したとするサンプルの中身に工作しなかったなどと考える方がどうかしてますよね。何れ、その問題は最後の課題となるでしょう。
(①の問題から順番に)SNPs解析結果は129が「僕のマウス」の片割れだと示唆しているにも拘わらず、その18番染色体にCAG-GFPが無いという結論は放医研の出したときのままになっている。そしてB6側の結論が間違っていたと判明した後に理研が129側を確認したか否かも知らされていない。
放医研の調査結果を若山さんが記者会見発表して後に、遠藤氏がそのデータを若山さんから貰って、放医研の出した15番にヘテロに4コピーのCAG-GFPがあると発表した結論は間違いであったと言った。放医研はCAG-GFPを探していたので、CAG-GFPのプロモーターの位置をプライマーで挟んで探していた。その時に、そのプライマーではアクロシン遺伝子の有る場所を検知してしまったのだという。支離滅裂の説明ですね。意味が分からん。
まず不可思議なのはFLSの15番に4コピー入っているとしたPCR産物は何かということです。どういうプライマーで挟んだのかということです。CAG-GFPの挿入された場所を探している。CAG人工遺伝子の並びそのものをプライマーにかけるということになる。
若山さんは自分のマウスはCAGだと説明していますね。だから放医研はCAGを探そうとした。そして順序はともかくとして「僕のマウス」には18番にホモでCAGがあるとした。若山さんは今まで自分のマウスのGFPがどこに入っているのか調べたことがなかったのでしょうかね。岡部マウスはもらった時に3番に入っていると聞いていたわけでしょ。岡部マウスは放医研では調べていませんよね。でも記者会見では3番だと答えている。普通自分のマウスのGFPがどこに入っているかは調べるよね。そもそもマウスの飼育はときどきコンタミが無いかどうかのSNPs検査にも出すものですよね。

FLS1-8の18番にはCAG-GFPが無いが、「僕のマウス」ESにはある。しかもホモで入っていると。
では、15番はどうか。FLS1-8にはある。しかも「僕のマウス」ESには無い。ところがこの15番は間違いだったという。では18番は正しいのか?
これが分からないわけです。桂報告書もGFPが何番染色体に入っているかをリスト上に記載していない。15番を間違えるような奴が18番は間違えなかったとどうしていえるでしょうか。そもそも探したプライマーを明らかにしていない。本当に両方とも同じプライマーで挟んだのかということすら疑われる。18番のCAGが正しいなら15番のCAGも正しいはずだ。アクロシン遺伝子はヘテロに4コピーなんてあり方はしないぞ。これは別のプライマーじゃないのか。
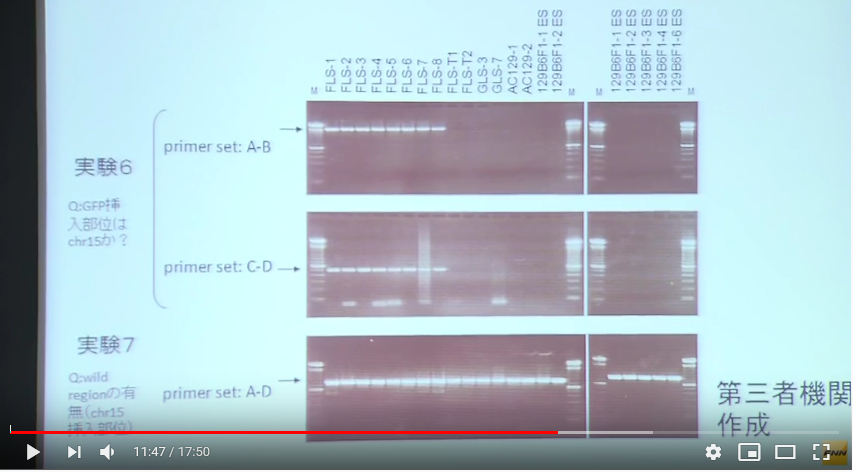
記者会見の画像ではプライマーのところが写されてないものもあって分かりにくいので、片瀬久美子氏のブログのを参考に以下に貼り付けて置こう。
上にどういうプライマーで挟んだのかが図示されている。コンストラクトというのは人工遺伝子のことで、無論これは若山さんがロックフェラー大学でB6にヴィルスヴェクターを使って遺伝子挿入したときのコンストラクトです。若山さんが行ったのですからその遺伝子構成は若山さんが知っていて、これを探すときに若山さんは放医研の知り合いに教えている。で、その末端につけられているポリAのあるあたりの部分にプライマーを設定した。要するに若山さんの人工遺伝子の尻尾を探したわけです。
実験3は18番染色体を調べた結果FLS1-8には存在せず、所謂「僕のマウス」シリーズにはあった。従って、若山さんの「僕のマウス」のGFP人工遺伝子は18番に入っていたことが分かったというものです。
このプライマーが分からないわけです。上の図のAの長さは286bpと書かれていて、プライマーの長さは通常20bp程度を前後に二か所設定するわけです。前はポリAのある場所ですが、後ろはコンストラクトの外に表示されている。でも、これは人工遺伝子全体が挿入された場所が分かっていませんからこの後ろのプライマーのDNAの並びは分からない筈ですね。プライマーが分かっていたら既にどこに有るかわかってるということですから調査する必要はない。
ど素人が常識的に若山さんの人工遺伝子を探そうとするとき、コンストラクト全体の遺伝子の並びは分かっていますから、頭の20bpと尻尾の20bpで挟みますね。そして染色体ごとに探すとまずGFPの存在している染色体番号は判明しますよね。もっとも頭の並びはプロモーターの並びなので内在性のプロモーター位置を掴んでしまうかもしれませんが、後ろの20bp以上並ぶポリAはDNAにはないので特定できそうですし、それも何らかの紛らわしいことがあるのなら、中にあるEGFP遺伝子の頭と尻尾に設定してもいいですね。要するに何番染色体に存在しているかということはそんなにむつかしくなさそうに思われる。
ところがこの実験3は妙なプライマー設定だと見受けられる。後ろのプライマーはコンストラクトの外に表示されてませんか。これって挿入位置が既に正確に分かっているコンストラクトの存在を再確認しているだけではないのか。
また、実験4は何をしようとしているのかがど素人には分からない。こちらは全体の長さが519bpです。そして図を見る限りはコンストラクトの少し前の位置から後ろは実験3のプライマーの後ろと同じようです。つまりこの位置の中にコンストラクトがあることがあらかじめ分かっているということです。そしてその区間にFLS1-8ではワイルドタイプの遺伝子が並んでいるのだということなのでしょうが、では「僕のマウス」シリーズではどうかというとバンドが何もない。つまりその中にコンストラクトが挿入されて長くなっているから519bp以上の長さになっているからこのバンドのラインには出て来ないんですね。
これって、どこに入っているかは事前に正確に分かっているということじゃないですか。若山さんは挿入位置を教えて、それがあるか無いかを放医研に確認させただけではないですか。
となると、岡部マウスの情報も分かった上で確認させているのではないかと疑われて来る。
岡部マウスは3番に入っていることは放医研で調べたことにはなってなくて、かつ記者会見では3番と若山さんが言ってますから、あらかじめ岡部さんから教えられている。でも、この調査ではアクロシンなんて思いもよらないという前提で放医研にCAG-GFPの在処を探させているということになっている。
マウスの15番には内在性のアクロシン遺伝子が存在しています。こちらは実験3,4の後です。FLSには18番にGFPが無かった。でも蛍光してますからどこかにはある。で染色体を虱潰しに調べたら15番で引っかかってきたという。変ですよね。これはCAGの遺伝子配列をプライマーに使っている。若山さんのコンストラクト全体の中でプライマー設定されている。15番にはEGFPはありません。因みにGFPとEGFPは同じです。蛍光発色の強化されたものです。
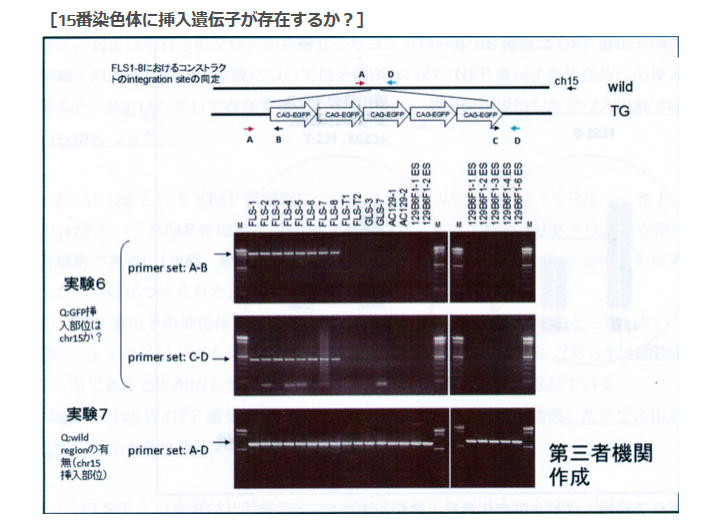
実験6は15番を調べたものと若山さんは記者会見で解説した。このPCR結果はあり得ないものです。GLSに無いのは当たり前ですが、そもそも15番にはGFPはない。それがFLS1-8には出ている。これは何なんでしょう。
そして実験7はA-Dで挟んでいる。これって若山さんのコンストラクトの中です。絶対に出ません。何なんでしょうか。
ひょっとして岡部マウスのコンストラクトを使ってないかと考えてみるとどうなりますかね。頭にアクロシンプロモーターがついている。すると15番にある内在性アクロシン遺伝子のプロモーターを掴むということはあるんですね。そして後ろのDの並びがあったかどうかでしょうね。
(どうしてこんなに分かりにくいのか)まずは実験3と4から確認していきましょう。見やすいように再掲します。
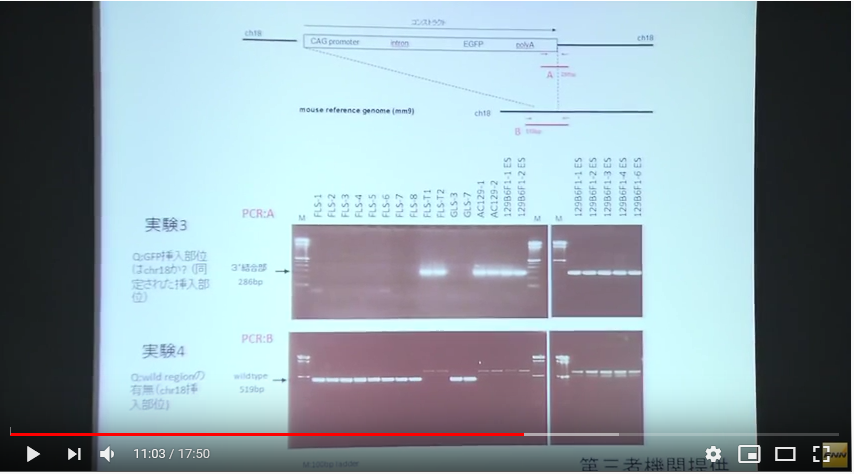
プライマーAはコンストラクトの後ろの方にポリAの並びがあって、その辺りからコンストラクトの末尾の少し後ろで挟んでいる。赤い矢印がそれですが、その長さは286bpと記載されている。そしてプライマーBはコンストラクトの少し前からプライマーAの後ろの位置と同じく、コンストラクトの少し後ろで挟んでいて、その長さは519bpと書かれていて、割り算するとプライマーAの長さは実寸でプライマーBの55%の長さです。この図でも赤の矢印とバーの長さと数値はそれを正しく示している。でも私は最初プライマーAはポリAの付近を挟んでいると思いましたが、他の人はどうなんでしょうかね。プライマーAのバーの長さと上の[CAG promoter---intron---EGFP---polyA]の概念図と比較すると尻尾の部分と誤解しますね。でもプライマーBはコンストラクトの長さを縮小していますよね。破線で縮小が示されている。プライマーの長さは数値で明確に書かれていますから、プライマーAはEGFPまで含んでいるんですね。全体の55% です。どうしてこんな紛らわしい表示にするのでしょうかね。全体を縮小しなかったらいいだけですね。何も面倒でないどころか、その方がシンプルで分かりやすい。縮小している意味が分かりませんよね。人が嘘をつこうとするときこういう姑息な手段を取りがちだということはまあ、警察のようにそれを仕事にしている人々だとか、そうでなくても比較的長く生きて経験豊富な人たちにとってはお見通しでしょうかね。しなくていいことをしている。分かりにくくするため以外には考えられない操作ですね。
コンストラクトの少しでも外側を使わなかったら実験4は出来ないというのは当たり前ですが、少し外側がプライマーとして知られているというのは事前にどこに自分のGFPが入っているかを知っているということです。
プライマーはAGCTの4種の塩基の並びの20桁程度を使います。4の20乗ですから組み合わせ数がほぼ1兆1千億程度になる。その20桁と同じ並びが他の場所にある確率は1兆1千億分の1ということです。マウスの全DNAの桁数は30億塩基程度ですから、余裕で同じ場所は無いと言っていいわけです。
[18番染色体に挿入遺伝子は存在するか?]ということを示している若山さんの記者会見での説明資料にある実験3は二つある前後のプライマーの後ろ側は若山さんのコンストラクトの外側にある。つまり挿入遺伝子は18番に有るということはあらかじめ分かっていて、かつ、どの場所にあるかまで正確にわかっていて、その確認をしているPCR実験です。もし場所が分かっていなくて分かっているのが若山さんのコンストラクトの遺伝子配列情報だけだったとしたら、その外側のプライマーはコンストラクト全体がどの染色体のどの場所に入ったのか分からないのですから作れません。
この実験はすべてが分かった後にその確認のために行われた実験だということになるわけです。[18番染色体に挿入遺伝子は存在するか?]という表題も又ミスリーディングなもので、このPCRで18番に有ることを探し当てたのではありません。既にどこに有るかわかっているものを確認しただけです。
実験4はその明らかな証拠で、プライマーBは若山さんのコントラストを内側に含んだ場所に設定してある。プライマーは業者に依頼して作ってもらうわけですが、どこに入っているか分かって無いと頼めません。これはプライマーの内側にワイルドタイプの遺伝子配列があるものがバンドに出ているものです。若山さんのコンストラクトが入っていたらその分長くなりますから、このバンド位置には出ないということです。これも分かったことを確認しているのであって、記者会見の席上で放医研とやらに調べてもらったなんて大げさなものではありませんね。このくらいは自分のところでできますね。どこに有るか分からないときは業種や専門機関に調べてもらう。高価な機械が必要です。この実験3と4はそのようなものでなく、放医研に頼んだとしたら結論が出た後の確認実験に過ぎません。或いはそもそも自分のマウスのGFP挿入位置なんて作成初期に専門業者に出して調べてもらっているはずのものなのですから、情報を全部与えてただ確認実験だけを第三者の立場だと称して行ってもらっただけのものかもしれない。そもそも放医研は研究所としては正式に依頼を受けたことは無いと言ってる。では依頼されたのは誰なんでしょうね。
次は実験6,7です。これも見やすいように再掲します。
この説明図を見て異様に感じるのは実験3、4では若山さんのコンストラクトを探している、もしくは確認しているはずなのに、ここではそのコンストラクトが示されていないということです。その代わりにCAG-GFPが5つ繋がった概念図が示されている。若山さんのコンストラクトの中のCAG-GFPは1つです。何を探しているのか、又、何を確認しているのでしょうかね。
[18番染色体に挿入遺伝子は存在するか?]というタイトルの実験は実質的には18番染色体のどの位置に有るかということまでが分かっている上での確認実験でした。ところがこの実験は違いますね。[15番染色体に挿入遺伝子は存在するか?]というタイトルで、こちらは本当にあるかないかも分かってない。探しているのは若山さんのコンストラクトではありませんね。検査されているサンプルのGFPがどこにあるのかを探している。若山さんのコンストラクトは「僕のマウス」ESの18番にあったがFLSの18番には無かった。しかし、常時蛍光はしているからどこかに何らかのCAG-GFPは入っている。ではどこにあるのかを調べようとしているんですが、CAG-GFPを探しているのですからCAG-GFPの構造の前後をプライマーに取ればいいんですよね。CAG-GFPの構造は分かっていますからその中でプライマー設定するとそれがPCRにかかってきますからCAGがあるということは分かる。でもこれではどこにあるのかは分からない。そもそも常時蛍光しているんですから有ることは分かっていますからそんな検査は無駄です。これを知るにはある一定区分で区切ったたくさんの種類のプライマーを作って、それをワイルドタイプと比較すればいいとど素人でも思いつきますね。GFPが挿入されている断片は長いので上に出てワイルドタイプは下に出る。二本出たらそこにある。ゲノムは全部読まれていて並びは分かっていますからあとは更に厳密に範囲を狭めて行ったらいいですね。
ゲノムウォークという手法があって、挿入遺伝子の構造は分かっているのですからどちらでも片方のプライマーは設計できるわけです。他方がどこに有るかわかってないんですからここにアダプターを付けて取り敢えずの場所の検討をつけた後にそのアダプターに反応した場所を新たなプライマーとする方法がある。以下はタカラバイオのゲノムウォーキングキッドの宣伝の中の図です。
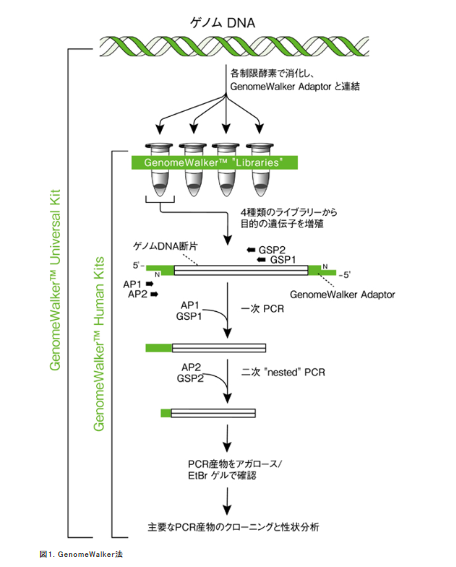
ど素人にはちょっとややこし過ぎて理解できませんが、まあ結論的に言えば場所の特定手法はちゃんとあるということです。業者に頼めば有料でやってもくれる。
ところが、放医研の第三者とやらは間違えたのです。マウスの15番染色体にはCAGは入っていなかったんです。ではこの15番染色体にヘテロで4コピーあったと報告したCAG-GFPは何であったのか。誰でも疑問に思いますね。竹市さんの疑問を貼り付けておきましょう。
>>
2014年7月22 日に訂正版を発表しました。
*ttp://www.riken.jp/pr/topics/2014/20140616_2/
2014年6月16日
CDB に保全されている STAP 関連細胞株に関する検証について
発生・再生科学総合研究センター
センター長 竹市雅俊
背景
STAP 論文では、脾臓から採取された免疫細胞が弱酸性にさらされることにより、多能
性を獲得することを報告した。STAP 細胞から増殖可能な細胞株として STAP 幹細胞が作
製された。STAP 論文に関する疑惑が明らかになった後、山梨大学若山教授は第三者研
究機関に STAP 幹細胞株の DNA 解析を依頼した。その情報は理研と共有され、理研は CDB
に保全されている STAP 関連幹細胞株の解析を進めてきた。
CDB 小保方研 STAP 幹細胞株の検討目的
CDB に保全されている STAP 関連細胞株(STAP 幹細胞、FI-幹細胞株)およびそれらから
作出されたキメラマウスの遺伝子情報を比較解析し、各細胞株間の遺伝子レベルの相違
と起源に関する客観的に検証可能なデータを得ることを目的とする。
CDB で保全されている小保方研細胞株の解析と結果
小保方研細胞株サンプルの遺伝子解析(遺伝的背景および CAG-GFP 挿入部位の確認)
• 小保方研細胞株サンプルのSTAP幹細胞6株に関して、遺伝的背景を6種のSNPマー カーを用いて検査したところ、以下の結果が得られた。GLS-1およびGLS-11につい ては核型の解析も行った。 1. FLS-3およびFLS-4: B6129F1: CAG-GFP, ♂ 2. GLS-1およびGLS-11: B6: oct4-GFP, ♂(核型の解析ではY染色体の一部に欠 失が見られる) 3. AC129-1およびAC129-2: 129B6F1: CAG-GFP ♂ (129 CAG-GFP マウス由来と されたが, 129B6F1由来であることが判明)。
• CDB では、若山氏から、STAP 幹細胞の CAG-GFP 遺伝子挿入位置の情報提供を受け、 上記 STAP 幹細胞株の CAG-GFP 遺伝子挿入部位を検証した。
STAP 幹細胞株 AC129-1 および AC129-2 は、18番染色体 GFP の挿入を持つ若山研 GFP マウスと同じ部位に、 1 コピーの CAG-GFP 遺伝子の挿入を持つことが判明した。かつ、相同染色体の両方に挿入されていることも若山研 GPF マウスと一致した。他方、FLS-3 および FLS-4 に関しては、15番染色体の片方の染色体に GFP 遺伝子が挿入されていることが判明した。また、CAG-GFP 遺伝子は複数コピーがタンデムに並んだ形で挿入されていた。これらの結果は若山研のサンプルの解析結果と一致した。
解析結果に対する見解
1.若山氏が提供されたとされる光るマウス(CAG-GFP 遺伝子保持マウス)から小保方氏が STAP 細胞を作成し、それを若山氏が受け取って樹立した STAP 幹細胞株に関して、保管されていたストックの解析から、CAG-GFP 遺伝子の挿入状況の違いにより、STAP 幹細胞は2種類の異なる遺伝子型のマウス由来であることがわかった。一方は、若山氏が提供した(CAG-GFP を18番染色体にホモで持つ)もの、もう一方は由来不明(CAG-GFP を15番染色体にヘテロで多コピー持つ)のものであった。
2.CAG-GFP を15番染色体にヘテロで持つマウスがどこ由来なのか、そのマウス個体が STAP 細胞から STAP 幹細胞が樹立された時期に若山研(あるいは小保方研)に生存個体として存在していたのかは不明であり、今後、さらなる検証を進める 誰でも15番にどうしてそんなものが見つかったのかを不思議に思いますよね。ここにはCAGはありません。にも関わらずどうして放医研の第三者といわれる人はここにCAGを4コピー見つけたのか。マウスの15番には本来の内在性アクロシン遺伝子があって、そのプロモーター領域もあります。仮に若山さんが岡部マウスのコンストラクトを聞いていてそれを教えていたのだとしましょう。ゲノムウォークするときに既知のプライマーがアクロシンプロモーターになっていた時には15番の内在性遺伝子領域をひっかけた可能性がありますね。でも、ここには4コピーなんて入っていません。4コピーあるのはCAGで3番に入っているのが岡部マウスです。間違えたにしても話がおかしすぎますよね。
(アクロシンプロモーター)放医研の第三者とやらはあくまでもCAGを探していたのだとしましょう。ゲノムウォーキング手法を使うとCAGが5個タンデムに並んだ断片が見つかったのだとしましょう。そして周辺を調べたらそこにアクロシンプロモーターが見つかったので15番だと思ったということですね。ところがそのアクロシンプロモーターは岡部マウスの人工遺伝子につけられていたアクロシンプロモーターだったということですね。
そして後に遠藤氏がそれに気が付いたという話になっているわけです。我々は誰が犯人か知っている。するとこのストーリーが作られたものだということはすぐわかる。その証拠が無いかということですね。
アクロシンマウスは若山さんは自分で維持飼育してますからね。「僕のマウス」B6でなければ、GOFのB6か、岡部マウスB6に決まっている。遠藤さんなんて出てこなくても可能性の中にはすぐ上がってくるものです。そもそも話が白々しいんですよね。あたかも自分は岡部マウスなんて考えもしてなかったなんて、最初のテラトーマからアクロシンが出でいて、このマウスが「僕のマウス」でないことは桂報告書の指摘するところです。FLSの時に「僕のマウス」を渡したということに実験ノートの証拠すらない。
岡部マウスの特徴は以下です。
>>
1)Acr-GFP/CAG-GFP 共挿入の位置、コピー数、周囲の塩基配列 表:STAP 幹細胞株一覧に挙げた 12 種類の幹細胞から STAP 幹細胞 FLS-T を除く 11 種 類の幹細胞株、それらの幹細胞が作製された 129 系統、および C57BL/6 系統の NGS によ る全ゲノム解析を行なった。その結果、Acr-GFP/CAG-GFP の共挿入は、STAP 幹細胞 FLS3、 FI 幹細胞 CTS1、そして ES 細胞 FES1 並びに FES2、ntES1 並びに ntES2, および 129/GFP ES の 7 株の第 3 染色体の同一部位に共通に存在することが判明した。また、Acr-GFP が第 3 染色体の片方にのみ挿入されていること(FISH により確認)、Acr-プロモーターの コピー数がどれも約 20 コピーであること、GFP 挿入部位を挟んで第 3 染色体の約 20kb の重複があることと、GFP 挿入部位に隣接して第 4 染色体 20kb 断片の逆向きの挿入があ ることも共通していることが判明した。これらの特徴は、2003 年に CDB 若山研が大阪大 学岡部研より導入した Acr-GFP/CAG-GFP マウスの特徴と完全に一致する。 (4P) ①3番染色体にAcr-GFP/CAG-GFP 共挿入がある。
②アクロシンプロモーターのコピー数は20コピー程度である。
③GFP挿入部位を挟んで約20kbの重複がある。
④GFP 挿入部位に隣接して第 4 染色体 20kb 断片の逆向きの挿入があ る。
⑤この特徴を共有しているのは2003年に岡部研より導入した岡部マウス、FLS3、CTS1、FES1、FES2、ntES-G1、ntES-G2、129/GFP ESである。
無論岡部マウスはホモ、その他はF1なのでヘテロに入っている。
ところで3番染色体には③④の二つの異常がある。しかし以下には欠失だと書かれていて、これが何なのかは上記引用箇所だけでは分からない。何度でも貼り付けましょう。
赤の四角は欠失と書かれていますよね。③④の中腹や交叉乗り換えとは違います。これって何でしょう。
>>
4)第 3 染色体と第 8 染色体の欠失変異
STAP関連11細胞株の全ゲノム解析から、第3染色体の5kbの欠失と第8染色体の17kb の欠失(第8染色体は129系統由来;第3染色体はB6系統由来)が上記STAP幹細胞FLS3、 FI 幹細胞 CTS1、および、ES 細胞 FES1 並びに 129/GFP ES だけに共通に存在することが 判明した。この2箇所の欠失は、STAP 幹細胞 FLS および FI 幹細胞 CTS の全ての株にも 共通に存在することがPCR産物の塩基配列決定により確認された。一方、この両欠失は、 市販の 129 の亜系である 129 x 1/SVJJmsSlc(SLC)と 129+Ter/SvJcl(CLEA)のいずれにも存在しない。また、この第 3 染色体の 5kb の欠失も、市販の B6 の亜系である C57BL/6JJmsSlc (SLC)、C57BL/6NCrSlc (SLC)、C57BL/6J (Charles River)、C57BL/6NCrl (Charles River)、C57BL/6JJcl (CLEA)、C57BL/6NJcl (CLEA)のいずれにも存在しない。 さらに、2010 年に若山研で受精卵凍結された Acr-GFP/CAG-GFP マウスにも存在しなかっ た。 もし、これらの細胞が論文に示されていた(129 x C57BL/6)F1 から作製された株で あるなら、これら 2 個所の欠失の両方、または片方が市販の 129 系統、C57BL/6 系統の いずれかに存在していなければならず、STAP 研究の行なわれた 2 年強という期間でこれ ら 2 個所の欠失が生ずることは考えにくい。従って、この結果は、これら 4 種類の細胞 が、論文に示されていた(129 x C57BL/6)F1 マウスから直接作製された株ではないこ とを明確に示している。(6P)8番は129側ですので今は論じない。B6の3番に別の欠失5kbがあるんですね。そしてそれを共有しているのは今度は、FLS3、CTS1、FES1、129/GFP ESです。岡部マウス、FES2、ntES-G1、ntES-G2には無い。
さて、いよいよ謎のキメラ子です。もう一度貼り付けます。
- 2019/11/07(木) 18:02:54|
- AC129
-
-
| コメント:3
(連絡)AC129の考察途中ですが、Ooboeさんから問い合わせがありましたので、返事を書きます。コメント欄では表示が制約されますのでここでやります。問い合わせは以下です。
>>
コメント
お久しぶりです
パートナーの手続き、進展中です。
桂調査委員の皆様にも検察申告書と
証拠資料集を添付して送付しました。
2019/11/07(木) 08:28:34 | URL | Ooboe #- [ 編集 ]
太田氏にも、送付する予定です。が
添付したい、桂報告と
論文との矛盾指摘必要箇所が
見つからないらしい。
教えてあげてください。
2019/11/07(木) 08:36:02 | URL | Ooboe #- [ 編集 ]
太田論文の何ページ目の何行目?
ですか?
2019/11/07(木) 09:04:31 | URL | Ooboe #- [ 編集 ] まず、桂報告書の方を提示しましょう。
>>
2−3.調査結果および評価
2−3−1.科学的検証等の結果から生じた新たな疑義の調査
2−3−1−1.STAP 関連の細胞株、キメラマウス、テラトーマに関する調査結果および評価
(a)調査に使用した細胞株(以下の(b)〜(d)などで使用した) 理研によりゲノム解析が行われた STAP 関連細胞株の一覧を下の表に示す。 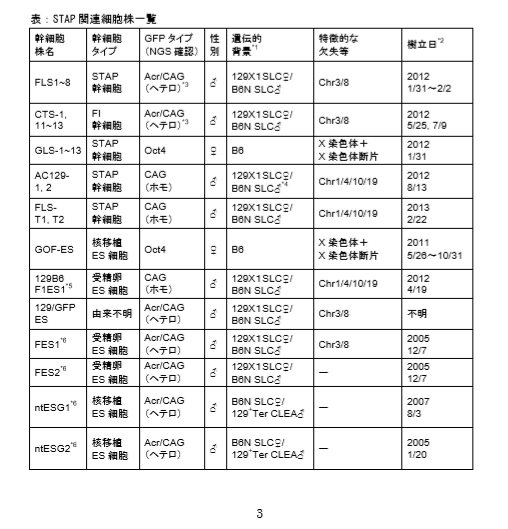 *1 親マウス系統と SNPs の比較解析により判定
*1 親マウス系統と SNPs の比較解析により判定
*2 細胞株培養開始日。ただし FES1 FES2 ntESG1 ntESG2 は凍結日
*3 作製者は 「CAG-GFP (ホモ)」と記載
*4 作製者は「129 CAG-GFP (ホモ) 」と記載
*5 129B6 F1ES1~6 が STAP 幹細胞に対する標準 ES 細胞として作製されたが、本調査に最も関わり が深いのは 129B6 F1ES1 である。
*6 正式名称は、 FES1:129B6GFP1 FES♂、 FES2:129B6GFP2 FES♂、ntESG1:129B6F1G1、ntESG2: 129B6F1G2この中で上段の 8 種類の細胞は、予備調査時に CDB 細胞リプログラミング研究ユニッ ト(以下「小保方研」という)、または山梨大学生命環境学部若山研究室(以下「山梨大 学若山研」という)に保存されていた STAP 幹細胞、FI 幹細胞、および関連する ES 細胞 である。また、STAP 幹細胞 FLS が、Acrosin プロモーター下に GFP を発現する Acr-GFP と、CAG プロモーター下に GFP を発現する CAG-GFP の共挿入を含むことが判明した後に、 過去に CDB ゲノム・リプログラミング研究チーム(以下「CDB 若山研」という)で作製さ れた Acr-GFP/CAG-GFP 共挿入 ES 細胞を取り寄せて解析したのが、下段の 4 種類の細胞で ある。ゲノム解析は、この他に STAP 細胞等の作製に用いられたマウス系統についても行 われた。「下段の4種類の細胞」がどこからどういう経路で取り寄せられたのかが書かれていないからOoboe さんたちが検察に調査依頼されているわけです。
これはチューブに入っている細胞でラベルが貼られている。その内容が注6に書かれているもので、
FES1:129B6GFP1 FES♂
FES2:129B6GFP2 FES♂
ntESG1:129B6F1G1
ntESG2: 129B6F1G2
コロンの右側が正式に記載されている文字です。ntESG1とntESG2は129がメスでB6がオスという表記です。ところがこれを調査チームが調べたら「B6N SLC♀/ 129+Ter CLEA♂ 」であった。この時点で既にラベル表記事項と中身が違っている。下段4種の細胞はどこかから取り寄せたと書いてありますね。その中の二つがラベル記載事項と中身が違っていたら、まず、その理由を調査しなければなりませんよね。そうでないと調べている4種すべてに関して細胞の信頼性がないじゃないですか。
ところが彼らは証拠能力を調査することなく残り二つの細胞の検査に入った。よく知られている通りです。
>>
他方、同じ Acr-GFP/CAG-GFP の挿入を持つ ES 細胞 ntESG1、および ntESG2 の X 染色体は C57BL/6 であることが判明したことから、調 査対象の STAP 幹細胞 FLS3、FI 幹細胞 CTS1 と性染色体の構成が異なるため、これ らは比較解析の対照から除外された。
いえいえ、サンプルのラベルと中身が違っていたら送られてきた先に問い合わせをしないといけません。なぜ違っているかの理由を聞いて、かつ残りの二つは本物なのかとも問い合わせないといけない。
このラベルには日付も入っている。凍結日ですがG1が2007/8/3、G2が2005/1/20です。これが太田さんの2005年論文と2008年論文に対応しているラベル表記だということです。
2005年論文は以前Ooboeさんとの連絡で知らせて、確かパートナー氏とネットカフェでダウンロード印刷してもらわれた経緯でしたね。それを相沢さんに見せたらよく調べてみるということで、相沢さんはもう一本の論文も見つけられた。
山梨大の若山研のホームページに関連論文の紹介があって、以前はこの二本の論文がありましたが、現在消されています。私は二本ともエクセルにコピーしていますから原文自体は持っていますが、この論文は今このブログではコピペすると書き込み禁止になります。たぶん若山さん側がコピペできないように処理登録したのだと思います。
ですから今原文を提示してここですとお知らせすることはできません。又論文にはページは打たれていません。
ただ、以前既に書いたように、グーグル翻訳したものを貼り付けると大丈夫のようです。
2005年論文は以下です。
>>
Generation of Normal Progeny by Intracytoplasmic Sperm Injection Following Grafting of Testicular Tissue from Cloned Mice That Died Postnatally
Biology of Reproduction, Volume 73, Issue 3, 1 September 2005, Pages 390–395, https://doi.org/10.1095/biolreprod.105.041673
Published:
1-Sep-05
Article history
Received:
8-Mar-05
Revision Received:
29-Mar-05
Accepted:
26-Apr-05細胞凍結は2005/1/20で論文の投稿は2005/3/5です。実験が終わってから論文を書くんですね。ここで使われたマウスは論文の中のMiceと書かれた項目の中にあります。原文を貼ると拒絶されますからグーグル翻訳を貼ります。そのままで手を入れません。
>>
未成熟雄B6C3F1、成体雌ICR、および成体雌BDF1マウスは、静岡研究所動物センター(浜松、日本)から購入しました。雌129 / Sv-terおよび雄ICRヌードマウスは、それぞれCLEA Japan、Inc.およびCharles River Japan、Inc.から購入しました。アクロシン/ eGFP(Acr3-EGFP)[34]およびpCX-eGFP [35]導入遺伝子(C57BL / 6TgN(acro / act-EGFP)OsbC3-N01-FJ002 [36]の両方を搭載した緑色蛍光タンパク質(GFP)トランスジェニックマウス)は、M。岡部博士(大阪大学、大阪、日本。すべての株指定は元の研究のものである)によって親切に提供されました。 GFP導入遺伝子を保持する129B6F1マウスを作成するために、雌129 / Sv-terマウスを雄のC57BL / 6 GFPトランスジェニックマウスと交配させ、GFP導入遺伝子のヘミ接合体であるこれらの交配の子孫を核移植のドナーとして使用しました。すべての動物実験は、実験動物の管理と使用のためのガイドに準拠し、理化学研究所神戸研究所の実験動物実験の組織委員会によって承認されました。これをそのままグーグル翻訳機に掛けて日本語から英語変換すると原文が見れます。
2008年論文は以下です。
>>
Increasing the Cell Number of Host Tetraploid Embryos Can Improve the Production of Mice Derived from Embryonic Stem Cells
Biology of Reproduction, Volume 79, Issue 3, 1 September 2008, Pages 486–492, https://doi.org/10.1095/biolreprod.107.067116
Published:
1-Sep-08
Article history
Received:
12-Dec-07
Revision Received:
9-Jan-08
Accepted:
25-Apr-08こちらはES Cell Linesの項目にあります。これもグーグル翻訳です。
>>
D3 ES細胞株は、もともとDoetschmanらによって確立されました。 [12]、およびGFPトランスジェニック細胞株(129SV(D3)-Tg(ACTB-EGFP)CZ-001-FM260Osb)は、pCAG-EGFP [10、11]を使用して確立され、岡部勝博士によって親切に提供されました。 (大阪大学、大阪、日本)。 E14 ES細胞株[7]は、1985年にエジンバラ(スコットランド)のマーティンフーパー博士によって同系交配マウス系統129 / Olaから派生し、ピーターモンバエルツ博士(ロックフェラー大学)から入手しました。 129B6F1G1 [13]およびBDmt2 [14]は、129B6F1バックグラウンドのSertoli細胞とGFPおよび雄BDF1マウスの尾端細胞をドナーとして使用して、当研究室で以前に確立された核移植由来ES(ntES)細胞株です。 それぞれ核移植。 GFPを発現するオスの129B6F1マウスは、メスの129 / SvマウスをオスのC57BL / 6 GFPトランスジェニックマウス(pCAG-EGFP [10、11]およびAcr3-EGFP [17]を使用して構築されたダブルトランスジェニックマウス系統[16])と交配させることにより作成しました。この中の129B6F1G1 [13]の脚注は以下です。
>>
13
Ohta
H
,
Wakayama
T
.
Generation of normal progeny by intracytoplasmic sperm injection following grafting of testicular tissue from cloned mice that died postnatally
.
Biol Reprod
2005
;
73
:
390
–
395
.
Google Scholar
CrossRef
Search ADS
PubMed追伸(AC129-8より)
アルイミオウジ説の話になったので序に書いて置きますが、彼はntESG1,G2のGをgenerationと解して、最初のntESがG1で、それを使ってキメラマウスを作って更にntES化したのがG2と推測している。そして実際に若山さんはこのやり方で20世まで作っていて別の論文があるわけです。でも太田さんのG1ははっきりしていて、2008年論文で2005年論文で作ったF1のntESを解凍して使った。その解凍した細胞を129B6F1G1と書いているんです。すると解凍する前の状態のラベルはどう書いてあるのかということになる。株分けした時に以前の分に129B6F1G2と書いたとしたらこれはgenerationの意味ではありません。ただ、日経サイエンスに太田さんは自分は2種類の受精卵ESと4種類のntESを作ったと証言している。太田さんに聞けばわかることで、彼には説明義務がありますよね。でも、チューブのラベルと中身が違っていることの説明は彼一人の判断では出来そうもありませんよね。
- 2019/11/07(木) 11:10:17|
- AC129
-
-
| コメント:5





























