AC129を巡る問題10からのつづきです。何かの文字と相性が悪いと書き込み禁止になるようです。L氏のコメントに対するこちらの応答に関する写真をここに貼っておきます。ここなら多分大丈夫ではないか。
自家蛍光というのはGFP以外の蛍光です。GFPの漏れ出しはGFPが光っている。この誤認に関して何か言ってる人もないですね。
>>
筆頭著者が不正をしないで誠実に真正なデータを残した場合でも、自家蛍光の誤認がそのままだと、Oct4-GFP偽陽性に基づいてキメラの実験に進んだと思われます。漏れ出しがあるのなら自家蛍光の誤認だと気づけないでしょう。デイリーも知らない事実でしょ。知らなければ漏れ出しは調べ方が適合的でなければ自家蛍光の中に含まれてしまう。だから考えが浅いと批判される。下の写真のOct4は内在性のOct4遺伝子が働いてOct4蛋白を作っているという証明で、Oct4-GFPが光ったからOct4遺伝子が働いたはずだという証明ではない。直接証明です。右端のGFPはローザクレですから細胞の由来を証明している。Oct4-GFPではないからOct4遺伝子発現の間接証明ではないが、実物が免染確認されているからそんな必要もない。以前L氏は両者の蛍光がずれていて同じ細胞ではないなどと言ってましたが、細胞塊はスライスされているからわずかにずれるんです。
この結果は丹羽さんのFigure3-bの結果と矛盾している。こんなにあっては変でしょ。同じ肝臓由来酸浴細胞塊です。要するに小保方細胞が何かということは完全にわかっているわけではないということです。こんなことはティシュー論文を読んだら最初からわかってることです。だからこそ研究しているんでしょ。小保方さんは正しく自分の細胞を再現している。キメラを捏造したのは若山さんです。最初は捏造のつもりでは無論なかったが、後の経緯でそうなってしまったということです。
(Ooboe さんからの連絡)旅行に出ている間に連絡がありましたので、ここで検討します。連絡事項は[Ooboeさん情報]に追加しています。ここにも再掲します。
>>
パートナーは、小保方stap研究を無きものにした画策を第一次、第二次、第三次の画策として分類した時系列説明資料を、検察に提出しています。
この中での犯罪事案は諸々存在していましたが、偽計業務妨害、背任、公務員法違反、公文書虚偽作成、告発幇助間接正罪などのうち、明確なもの、2件にしぼって、告発手続きでなく、申告書提出の手続きをしているところです。
検察からの文書も全てではありませんが、一部公開する予定です。担当官は公開okとのことです。
2019/11/27(水) 13:30:12 Ooboe
第一次共謀画策は、2014年3月~6月16日
第二次共謀画策は、6月25日~12月26日
第三次共謀画策を、2015年1月~6月
と大きく分類しています。
この経緯の中で、分水嶺時点を6月5日として検察に資料を集中して説明しています。
そのパートナー証拠の更なる裏付けをお願いしていますが、頑張れFBでは、詳細は控えるようです。
全経緯を通じての、解明ポイントを6月5日の理研本部会議時点として捉えると、6月25日以後の第二次共謀画策や第三次共謀画策の経緯の本質が浮上してくるので、犯罪事案の、間接的ではあるが、背景として、その内容を検察に理解して貰うのに焦点をあてたとのことです。
この詳細は、ブログ開設で展開したいそうですが、検察には、この首謀者と共謀者を証拠をもって特定して提示しているとのことです。
この首謀者については、居士さんの考察とは異なるものとなっています。
第一次画策以前の2011年から2015年の第三次画策の全経緯を通じて一貫した首謀者としてパートナーは検察に提示して説明しています。
抽象的で、ごめんなさい。
2019/11/28(木) 00:33:12 Ooboe ①小保方stap研究を無きものにした画策
②共謀画策は三次にわたる
第一次共謀画策は、2014年3月~6月16日
第二次共謀画策は、6月25日~12月26日
第三次共謀画策を、2015年1月~6月
③分水嶺時点は6月5日の理研本部会議時点
④犯罪事案は以下の内の二件に絞って申告書提出手続き中
偽計業務妨害
背任
公務員法違反
公文書虚偽作成
告発幇助間接正罪、等
纏めると以上でしょうか。6月5日の理研本部会議時点がエポックであるということは会議に参加していた人たちの中に首謀者、もしくは首謀者にそそのかされている人が潜んでいるということになりますね。我々の今までの交換情報の中では松崎氏ということになると思います。
「この首謀者については、居士さんの考察とは異なるものとなっています。」の意味が二通り考えられるので、このことに関して少し述べておきましょう。
私は"犯人"は若山さんと言ってます。この意味は明確で、キメラができたのは若山さんのntES化手技の結果で、論文に書かれた小保方細胞の自然の結果でもなければ、小保方さんのESコンタミ捏造でもないという意味です。その意味で、私は犯人は若山さんだと言ってる。
対して、まず最初に、パートナー氏の(Ooboeさん、和モガさん、Ts.Markerさん、学さん、アルイミオウジさんをも含むかもしれませんが)推測の前提として明確に私のストーリーと対立しているのは首謀者ではなくて、論文通りにキメラのできているSTAP細胞があるか無いかという点です。
①論文通りのSTAP細胞由来キメラがある。
②小保方さんがESによって若山さんに捏造させたキメラがある。
③若山さんが小保方細胞核をntES化したものからのキメラがある。
パートナー氏の推理は①です。私は③です。桂報告書は②です。
②はありません。パートナー氏も私も否定済みですね。ため息ブログや雑談コーナーももはや小保方さんの論文不正を言い立てるばかりで、なぜキメラができたかに関しては口を閉ざし始めていますね。あそこにたむろしているのは小保方捏造を騒ぎ立てたマスコミ関係のスピン屋たちばかりです。言い訳集団です。医学界関係の論文不正というのは昔から有名でね。医者が自分の箔付けのために博士号を欲しがるので、一回限りの論文ということで審査や内容はユルユルで、不正は昔からまかり通っている。細胞生物学会にもはびこっているんですね。だいたい学会というのは利権団体ですからね。言うまでも無いが立派な医者、立派な研究者も昔から少数ながら常に存在している。そうでないと社会は維持できない。
小保方さんの不正の中で一番重大なのはメチル化検証です。これは若山さんがキメラを作る前からもちゃもちゃいじっていてプログレスレポートでいい加減な報告をしている。これは若山さんに誤解を与えたかもしれません。ただし、三誌論文にこのデータを使ったというのはキメラが出来ているんですから、若山さんが誘因になっている不正です。無論、不正は不正で、そもそも研究というのは真実を知るために実験しているのであって、実験事実に忠実でないものは、研究姿勢に既に問題がある。モラルなんて問題じゃない。ましてお作法なんて落第生に言う低レヴェルの話だ。意図しない結果を実験が教えてくれているのにそれを無視するなんて研究者じゃない。心が鍛えられていない。もしくはふんどしが緩んでいる。
①は再現実験ではできなかったので論文発表が早すぎるということは明確にされたが、あの再現実験は制約がありすぎますし、何をしたかに関して、隠されていることが多すぎる。特に三胚葉分化実験をしたかしなかったかに関してはとても深い疑義がある。その意味でできないということが証明されたとは言えません。でも、あり得た場合になぜ、理研が世界に向かって記者会見発表までしたものを後に無かったことにしたのかの説明が必要です。誰が、できているものをできていないことにして隠そうとするのかを考えないといけない。
この結果は世界的に大きな利権になるから理研が隠そうとするということはあり得ませんね。世界に向かって記者会見したのは理研です。
もうしてしまってから、初めてそれを知って、理研に対して隠せと命令できるとしたらもっと上の組織で、官僚組織ということになるが、これもないですね。笹井さんは文科省との折衝役で事前に文科省に説明しています。小保方さんをつれて行ったこともあると手記には書かれている。記者会見発表前には文科省からの天下り役員も知っていて連絡していますね。こういうのはないです。
「小保方stap研究を無きものにした画策」という言葉の中にSTAP研究と書かれているが、この研究はキメラができたことを事実として含んでいるものなのか、それとも理研以前の段階、つまりティシュー論文の研究までを意味しているのが曖昧です。
後者であっても現実的には小保方さんの博士号を剥奪し、ヴァカンティと小島両氏の地位を失わせたのですから、"無きものとした画策"ということはできます。どちらなのか、どちらもなのか、明確にしたいものです。
ただ、この課題は無数にある科学的課題の中の一つに過ぎません。時がたてばこの研究に興味をもってやり直す人は出てくるでしょうから、隠さなければならないほどの利権を生み出す課題でもないですね。利権になるのは臓器にまで作れた場合で、ESですらとんでもなく遠いのですから、こういう基礎研究を隠すなんてこともありません。笹井さんを失ってもっと遠くなってしまったとも言えますね。でも、弟子が続くでしょう。
[①論文通りのSTAP細胞由来キメラがある。]のに、なぜ隠そうと画策するのか。大した利権でないのに、隠そうとした動機は、だから[論文通りのSTAP細胞由来キメラがない]からだと、私は考えたんです。
私の方に来るためにあなた方をそちらに引き留めている理由をちゃんと書かないといけない。逆にその理由に私が納得したら、私の方からあなた方の方に行くでしょう。
私はあなた方が意図する意味での理研内の首謀者としては松崎氏であると考えている。笹井さんに対する内部権力争いにこの問題を利用して、報告書の基礎になる実験結果を提出した。ただし、ここには早く収拾しろという文科省の圧力もあって、利害が一致したということはありそうですね。もう一人松崎氏以外にGDがいますね。そちらですかね。そこは私は分からない。
無論、私は松崎氏を誘導したのは若山さんだと思っています。嘘をついて騙している。松崎氏はそれを信じて実験を行っている。ESコンタミ捏造だという結論を想定して実験を行っているんです。報告書が若山さんを全く容疑者として扱ってないことは自明ですね。全く客観的な調査ではない。
私のストーリーが正しいとして、後の若山さんの行動から人々は若山さんを疑えたかと考えるべきです。結構上手に騙しているんです。そもそも科学者はこんなことで人を疑いませんからね。私のストーリーに思い至れない人々は小保方さんのESコンタミとしか思えませんよ。たくさんそうでない事実を見ているんですけどね。若山さんがESコンタミ捏造なんてするわけはありませんからね。すると意図しない経緯でこうなったという事情は気づいていませんから調査できません。こういうことにはプロの捜査に慣れている警察を入れないといけないんです。業界の自主運営ではいけません。欧米は皆そうなってる。
(学さんの誤解)序なので、2019/11/28付の学さんの考察記事に関してコメントしておきましょう。誤解がありますね。以下です。
>>
年数が経つと、1塩基変異が積み重なってきて、コロニーごとのマウスのSNP(1塩基単位の突然変異を含む)が、離れてくる現象について議論してます。129であれ、B6であれ、その近交系マウスに特異的SNPsは100万か所単位で存在しています。調べているのはその中の一部だけです。まして、登録されていない新たにできたSNPsなんて調べられてはいません。
Ts.Marker さんのブログの2015/12/31の記事に以下のようにある。
>>
------ 追記 6/30 -----
マウスのSNPsは1000万といわれていますが、調査報告ではB6と129で違う300万を使用している。(あのねさんへの返事)学さんのブログにあのねさんがコメントされているのに気づきました。以下です。
>>
あのね
このコメントを参考に考えて見ましょう。一言さんもブログで、なぜRosa?と書かれたようにブログで追加ご考察して下さい。
>5603. L
2019年11月17日 06:58
モンキーさんのコメントに納得できないのは、あまりに笹井先生に失礼な推測だと思うからです。笹井先生がそんないい加減な形でメソッドを書くとは考え難いです。なので、筆頭著者が書いた(あるいは筆頭著者からのインプットで笹井先生が書いた)と私は推測します。この場合は、まず、筆頭著者に誰かからRosaマウスについてのインプットがある必要があると思いませんか? だとすると、どのような意図でインプットがあったのか、疑問に思いませんか? 謎でしょ?
Rosa26の遺伝子座にGFPを挿入したマウスは、理研CDB相澤先生が動物資源開発センターの室長兼任で当時、確立してして論文発表しています。アブストを貼り付けますので日付に注目して下さい。2011年7月前後の時期に若山研究室ではどのようなSTAP実験をしていたか想像しましょう。
丁度、若山研でGOF-ESを作って、後に2つの近交系129マウスとB6マウスを交配させてF1マウスを作ろうとしていた時期だと思います。この時期はすでにCDB内部で129 carrying Rosa-26GFPマウス完成の案内通達があったはずで、研究所内で都合の良い129GFPマウスが配布されているのに、市販の129マウスだけを買う人はいません。若山氏はこのマウスも使ったと思います。後にこのマウスを調べると困る流れになります。FES1の由来のマウスに合わなくなるからです。そしてそんなマウスは無いことで済ませて、遺伝子配列は別口ですでに登録されているのにゲノムの比較調査されていません。
http://www.cdb.riken.jp/jp/04_news/articles/11/111212_liveimagingmice.html
独立行政法人 理化学研究所 神戸研究所 発生・再生科学総合研究センター
2011年12月12日 「ライブイメージングに適した新たな蛍光マウスを開発」
発生過程において組織や器官が形成されていく仕組みを知るためには、個々の細胞の振る舞いを調べる必要がある。蛍光ライブイメージングは、生きたままの胚において特定の細胞や細胞内の構造を可視化し、その動きを追跡することができる技術である。マウスにおいても蛍光ライブイメージングが可能だが、多くの場合、蛍光遺伝子を染色体にランダムに導入しているため、「←ここ重要」発現部位を厳密に制御できない、発現量にバラツキがある、複数の構造を同時に標識できないなどの問題を抱えていた。理研CDBの動物資源開発室(相澤慎一室長)は、核や細胞膜など7種類の細胞内小器官を条件特異的に蛍光標識できる12系統のマウスを開発した。ライブイメージングに適した十分な蛍光シグナルが得られ、また、二重標識も可能であることが確認された。既に汎用されているCre-loxPシステムを発現制御系に用いているため、容易に発現部位を限定できる。この研究成果はGENESIS 誌の7月号に掲載され、同室はこれらのマウスの配布を開始している。
これが129/Sv carrying Rosa26-GFPマウス。ところが、桂報告書10P
「…なお、Article のメソッドに、129/Sv carrying Rosa26-gfp からキメラ寄与能を有する STAP 幹細胞が樹立された、との記述があるが、129/Sv carrying Rosa26-gfp マウスは理研 CDB に導入された記録や飼育記録はないことから、これは誤記と考えられ、若山氏の説明によればここで言及された STAP 幹細胞は AC129である可能性が高い。」
CDBに導入された記録や飼育記録はないなんて、よくも平然と言えたものですね。「これは誤記と考えられ」と含みを残して「誤記である」となぜ断言できないのですかね。少なくとも小保方さんが笹井先生らにSTAP細胞のライブイメージングで見せて「良く光っているね」と感想をもらしたマウスはRosa26-GFPマウス(若山研B6と交配F1を含む)のSTAP細胞であるほうが自然かなと思います。自己保身で調査に率先して協力した=小保方研に都合の良い細胞だけを残し、泥棒細胞を抱えたオフサイド若山さんの言質を取った調査委員会は市販の129マウスだと信じた。確証バイアスは恐ろしい。CDBで配布されたマウスも記録なしとなる。STAP論文が取り下げられて、なおかつ追撃でSTAP現象すら否定なら「嘘も方便」とはこのことですね。
調査報告書でも指摘されていたように、若山さんのマウスコロニーの管理の無頓着さについては、「あの日」の記述から考えられる考察案件を持っており、追ってそもそも論から後にコメントします。
2019/11/29 URL 編集この件は既に考察済みなんですが、あれこれと何度も考え続けていますから、人にはなかなか分かりません。再説しますと、この相沢さんのマウスは6種の色違いの蛍光遺伝子とクレロックスシステムを使って最終的に12種類の同時識別可能なレポーター遺伝子を造れたということです。その時に使われたマウスの背景はここには書かれていません。従って、129/Sv carrying Rosa26-gfp マウスは無いと言われたらそのまま受け取るか、無いという証拠を出せというよりない。誰もその試料を開示請求していません。
私が今考えているのは誰かが小保方さんに129/Sv carrying Rosa26-gfp だと言ったから、又誰かがキメラを作ったと言った、もしくは言ったと誤解したから、彼女は論文にそう書いているということです。
そもそもなぜ若山さんがこんな時期に背景マウス種の違いによるSTAP細胞の樹立確認をしたのか、しかも後にキメラは作らなかったと言っていて、確認目的ならキメラまで作ってみないと変なので、これは目的が違っていて、TCR再構成結果に対する別の関心から小保方さんに129のSTAP細胞を作らせたのではないかという可能性を今、考えているんです。
(和モガ氏のヘテロプラズミー)Ts.Marker さんがいろいろと調べていましたが、最終的にNOD / ShiLtJまで突き止めてその後の進展が無かった。このマウスは免疫不全マウスなので、このようなマウスはクローン作製には使われない筈なので変だなと思考停止していました。この件に関して和モガさんも考察を続けられていたのはずっと読んでいましたが、今又読み直して読み落としに気づきました。以下のように書いている。
>>
NOD / ShiLtJマウスを提供しているCharles River社のホームページにはNOD / ShiLtJマウスは非近交系のICRマウスが起源であると書かれている。ICRは若山研でクローン胚を作るときのマウスです。ただ、Ts.Marker さんはICRを調べてない。
>>
-------------- 12/7 調べ -----------------------
Mouse strain assembly hub - May 3, 2017 より
DBA/2J T (https://www.ncbi.nlm.nih.gov/nuccore/CM004183.1)
129S1/SvImJ T (https://www.ncbi.nlm.nih.gov/nuccore/CM004179.1)
BALB/cJ T (https://www.ncbi.nlm.nih.gov/nuccore/CM004214.1)
C57BL/6NJ T (https://www.ncbi.nlm.nih.gov/nuccore/CM004277.1)
NOD/ShiLtJ A (https://www.ncbi.nlm.nih.gov/nuccore/CM004185.1)和モガさんの整理しているものもここに貼り付けて置きましょう。分かりやすいです。
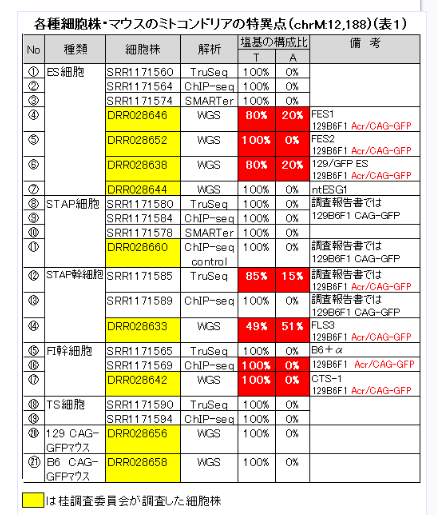
ICRのデータがAであれば、私の小保方細胞核使用ntES論にとっても強力な傍証になるでしょうね。ただし、ヘテロプラズミーは必ず起こるというものでもないんですね。
(学さんの誤解2)まだ学さんの誤解が続いているようです。ため息氏とのやり取りですが、ため息氏の認識が正しく、学さんは勘違いしている。これで双方で悪口の言い合いをするので、小保方さん擁護の学さんの旗色が悪くなるから、世間はスピン屋たちが正しいと誤解する。私はこれをスピン屋同士の芝居だと見做したわけですけど、Ooboe さんはそうではないとおっしゃるので、学さんの誤解だとして指摘し続けなければならないようです。
>>
F1では、129とB6がランダムに混じりますので、子どもごとに違う遺伝子構造ですが、129B6ES1~6の例でもわかるように、両親がそれぞれ、同じ遺伝子構造なので、その子どもごとに129とB6が混じり方が違っても、それでも、子どもの遺伝子の類似性が高いことがわかります。説明の仕方が悪いんですね。「F1では、129とB6がランダムに混じります」というのは近交系が崩れている時の話ですよね。先に、完全な近交系ではランダムには混じらないという原理から順番に書かないから支離滅裂になるんですね。完全な近交系が保たれていたら「子どもごとに違う遺伝子構造」にはならない。何匹生まれても全部同じ遺伝子構造のF1になる。
それを説明した後に、マウスコンタミがあって近交系が崩れている場合は「F1では、129とB6がランダムに混じりますので、子どもごとに違う遺伝子構造」になると説明したらいいんです。
最初に説明すべきことを誰でも知っていることという前提で端折るだけなら、少しずつ違うのは当然だということで分かる人には誤解が無いんですが、逆になぜ少ししか違わないのかという説明として、後から、誰でも知っていることとして端折ったはずの説明を加えるから、あなた、どういう理解をしているのと、分かる人から反論されることになる。
>>
F1の場合は、それぞれ近交系で、129コロニー、B6コロニーでは、ほぼ同一遺伝子状態であっても、交配してしまうと、できてくるF1マウスは、それぞれ129とB6の混じり方が異なってくる。ここも、「交配してしまうと、できてくるF1マウスは、それぞれ129とB6の混じり方が異なってくる。」のは近交系が崩れている場合ですから、前の文章に「F1の場合は、それぞれ近交系で、129コロニー、B6コロニーでは、ほぼ同一遺伝子状態であっても」という説明を置くと前後の文章が矛盾してしまう。「ほぼ同一遺伝子状態」が「同一」だと言ってるのか「同一でない」といってるのかというと、「も」が入っていることによって「同一」だと言ってることになるんです。作文の仕方が間違ってる。日本語の「も」の使い方の間違いです。学さんは日本人ではないのかな。
>>
2019年12月2日 4:12 PM
>学とみ子は両親がヘテロザイゴート のとき作成される配偶子は均一にならないからその子供同士のSNPは一致しないのがわからない?
近交系というのは兄弟同士等を掛け合わせて、すべての遺伝子(DNA)をホモザイゴートにしたマウス のことでしょ?ホモザイゴートだと減数分裂にのとき、交叉が生じても配偶子の遺伝子構成は変わらないから兄弟のSNPsはほぼ一致する。ため息さんの頭の中には、過去に129とB6が混じってしまった話と、自然に生じる塩基変化で遺伝子型が変化していく話とが区別つかず、混乱しているようです。
ホモザイゴートとは、129のみで維持、B6のみで維持の場合ですよ。事故的に他のマウスが混じっても、又、ホモザイゴートのコロニー内で、マウスの遺伝子は変化していきますよ。①完全に近交系マウス同士のF1は何匹生まれても両親から半々に遺伝子を受け取るので完全に同じ遺伝子構成になる。子供同士でわずかずつ違うということはない。
ため息氏は近交系の基本認識を言ってるだけです。野生のマウスは両親の遺伝子を受け継いでいるので減数分裂の時にできる配偶子は遺伝子の並びが引き継いだ両親の遺伝子に分かれるので2種できる。でも、近交系のマウスはその両親が同じだから減数分裂しても全く同じ配偶子ができる。だから129とB6を掛け合わせた子は何匹生まれても遺伝子構成が全く同じ混血の子供たちが出来るんです。混血ではあるが子供同士で遺伝子の並びに違いがあるということはないんです。この時点でため息氏は何も間違ってない。
②マウスコンタミがあって近交系が崩れていたらどうなるか。
既に論じたように、若山研でのマウスコンタミには二つの要素があって、129に関してはそもそもジャクソン研究所で古くにB6の飛び込みがあって気づかれていなかった。
もう一つは、若山研で自家繁殖させている間に生じたマウスコンタミがある。
SNPsに関しては前回指摘したように、調査されているSNPsはそれぞれの近交系マウスの特異的SNPsとして登録公開されている1千万個程度の中の、特に、今回では129とB6で違っているSNPsの3百万個だとTs.Marker さんが書いていると指摘した。
SNPsはマウスを飼育していると生殖細胞にも突然変異が起きてきますから、最初に登録されていた1千万個に対して継代ごとに何個か、何十個かずつ増えてくる。でもこれは最初の登録されている場所とは違います。1千万個に対して有意なだけ増えると別種の近交系マウスとして新たに特異的SNPsが登録されます。それまでは変更ありません。学さんは新たに増えたものを調べていると思われているが、そんなところは調べられてない。
次にSTRに関してはFES1とFES2で違っていたからFLS3はFES1と同じだと東大グループと遠藤氏が結論したと日経サイエンスに書かれている。我々は無論若山さんがFES1のチューブにFLS3を入れたからだと思ってますが、それはともかくとして、同時期に同じ近交系マウス同士の親を交配して作ったらしきFES1とFES2のSTRが違ってるなんてありえないと知らねばなりませんね。
FES1とFES2のSTRが違いうるという説明を太田さんにして欲しいね。どんな交配したの。③培養変異の存在。
これは体細胞分裂ですからね。別の話です。
さて、学さんの「も」の使い方は変なんです。「ホモザイゴートとは、129のみで維持、B6のみで維持の場合ですよ。」と書かれているのは近交系なら当たり前のことです。そしてこのことをため息氏が言ってる。言ってることが同じならここで又確認する必要は無いでしょうよ。そこにどうして、「事故的に他のマウスが混じっても、又、ホモザイゴートのコロニー内で、マウスの遺伝子は変化していきますよ。」という訳の分からない文章が続くのか。
「事故的に他のマウスが混じっ」たら、既に近交系マウスではなくなっている。それはここには引用してないがため息氏は分かっている。それは向こうのブログに書いてありますね。ここで学さんが引用しているため息氏の文章の近交系マウスの認識は正しいんでしょう。そういう場合はまず正しいと認めた上で、私が言ってるのはマウスコンタミのことだと言えばいいだけでしょ。私の言いたいことはそこでは無いと。
でも、「事故的に他のマウスが混じっても、」という文章はどこに落ちていくんでしょうかね。事故的に混じったら近交系ではなくなる。はい。で、「又、ホモザイゴートのコロニー内で、マウスの遺伝子は変化していきますよ。」とは何を受けている文章なのでしょうか。これって「事故的に他のマウスが混じったら、又、ホモザイゴートのコロニー内で、マウスの遺伝子は変化していきますよ。」の間違いじゃないの。「も」が入ると意味が変わる。日本人じゃないんですかね。外国人ならこういう間違いは理解できますけどね。
学(ガク)さんって本名なら珍しいお名前ですが、ひょっとしてシナ系の帰化人の方ですかね。もしそういう帰化人1世のかたであったら、このことはとても理解可能ですね。そしてそういう配慮で読むことができる。
「も」の使い方の指摘は2度目です。なんとなくそうではないかと思えてくる。
ため息氏のスピンはそんなところにはないので、こういうやり取りをされて、本質的でないところで学さんがやり込められると、小保方さん擁護派は困るんですがね。
(アルイミオウジ氏の最近の動向)アルイミオウジ氏を常にチェックしているのは最近では私くらいのものではなかろうか。
上の学さんの相手が自分のブログのコメント欄に書いているのが以下です。
>>
sigh
2019年11月22日 9:21 AM
>学とみ子
擁護の阿塁未央児さんも「学さんは何が言いたいのだろうか? FES1とFES2で雄親のコロニーが違うという示唆が、桂報告書のどこにあるのだろうか?」と呟いていますよ。
何が言いたいの分かっている方はいないようです。説明したらどうでしょ?ここで紹介されているアルイミオウジ氏のツイートが以下です。
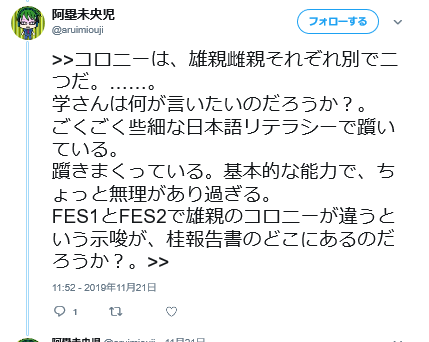
学さんが言ってるのは上述したように同じコロニーから作られた筈のFES1とFES2のSTRが違ってるのはなぜかということでしょうね。
このこととは別に最近アルイミオウジ氏が面白いことを発言している。以下です。
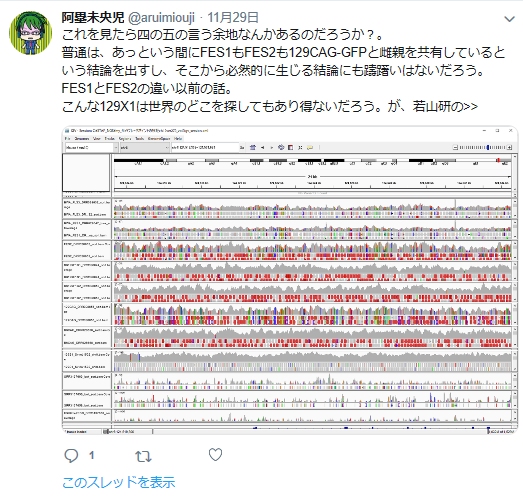
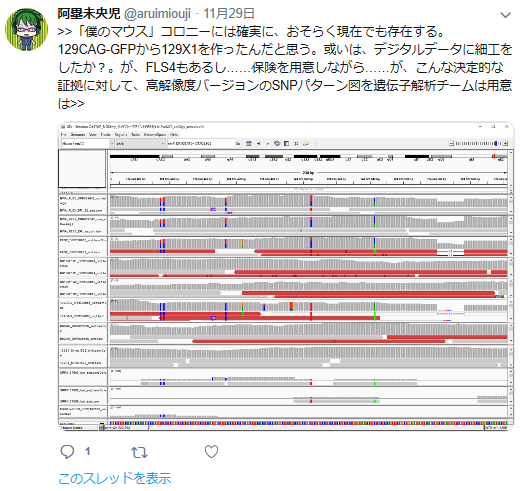
私の疑義していたことですね。
>>
AC129を巡る問題8
(残された問題の整理)
さて、問題の数も少なくなってきましたね。ただし、解の困難さは増してきました。ここらで残されている問題を整理しておきましょう。
①FLSは「僕のマウス」を渡したと一方的に証拠の提出もなく若山さんが主張しているが、実際の検査で出て来たのはAcr-CAGのB6(岡部マウス)と市販のGFP無しの129X1/SvのF1だという報告書の調査結果となっている。しかし、129側の18番染色体に1コピーのCAG-GFPが入っているのではないかという疑義がでた。なぜなら、SNPs解析結果がこの129は「僕のマウス」の片親であることを示唆しているからだ。最初に放医研がFLSを検査した時、15番にヘテロに4コピーのCAGがあり、18番にはGFPが無かったと発表していたが、後に15番のGFPが否定されたが18番に関しては何も触れられなかった。又間違いの原因もちゃんと説明されていない。内在性アクロシン遺伝子を間違えたのならホモだと思うはずでヘテロに間違うというのは考えにくい。調査自体がとてもあやしい。すると18番に無かったというのは本当なのかという疑義が残るということです。理研のチームはGFPを自分でちゃんと確認したかということです。この説明も無い。アルイミオウジ氏は学さん以上に説明能力の欠如している人なのでどういうことをしているのかということがちゃんと確認できるまでは何とも言えませんが、129CAG-GFPマウスから129X1を作ったという説明は聞き捨てならないところです。これは129X1に戻し交配したということですが、129X1があるのに戻し交配してGFPを消すという意味が分かりません。これはGFPの有無を調べればいいだけのはずですが、GFPは無かったのでしょうか。ちょっと明確でないところです。ただ、129にB6のコンタミしているSNPs分布が「僕のマウス」の片親である129と同じだという分析結果は私と同じですね。
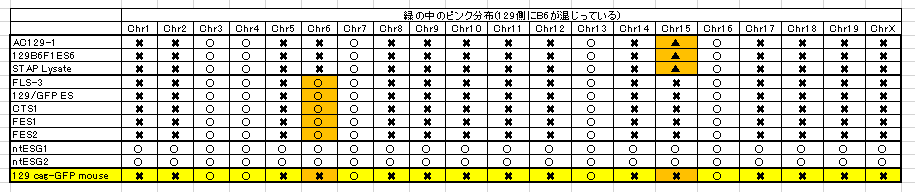
また最近彼がやってるのはSNPsでなくてSNVsを見ているらしい。頻度が1%以上ならSNPであるが、それ以下まで入れると無数にあることになる。何をやってるのかちっともわからない。以下も、勘違いということでいいんだよな。
 (学さんの誤解3)
(学さんの誤解3)学さんが何を誤解されているのかが段々はっきりしてきましたね。
>>
近交系マウス由来同士に、予期せぬ交雑がおきると、別の系統のSNPが、マウスに入ります。
さらに、交雑が進むと、別の系統のSNPをホモでもってしまうことがあります。
かつて混じったというと証拠でしょうが、近交系コロニーに戻して年数が経つと、次第に別の系統由来SNP変化が少なくなり、ワトソンさんらは、薄まると表現されたと思います。
以前に、専門家の方が、ヘテロはホモに収束するとか書ていたのを見ましたが、細胞は独自に、SNPの修復を行うと考えて良いのかな?、ここは確かでないです。近交系マウスは基本自家繁殖させてはいけないです。マウス供給業者との契約でそうなってるはずです。
近交系マウスは兄妹交配を続けます。親にはかけ戻しません。親の管理ケージと生まれた子供たちの管理ケージは分けられています。
兄妹交配した子供は親たちよりホモ率が高いんです、その原理を使って20継代以上させて98%以上のホモ率になったものを近交系マウスと称して業者が販売している。
その後、買ったマウスの雌雄を交配させて維持繁殖させるときに、業者と同じように兄妹交配管理を続けているとホモ率はどんどん上がっていきます。以前そのデータを貼り付けましたね。
でも、買ったマウスをどのような管理をするのかは研究者たちの自由ですが、自分で繁殖させたものを使って論文を書くときに購入業者の名を使うと違法になりますね。論文にマウスの業者を特定させたいときは買ったものそのものを使わないと業者は責任取れません。今回みたいにこれだけのマウスコンタミがあると研究目的によっては責任問題になりかねない。間違った取説で使用されたものにメーカーは責任取れません。
若山さんのところではいろんなマウスを自家繁殖させていました。クローンの研究者ですので、作られたキメラの貢献度を見るためにGFPが入っていればいいんですね。基本自分たちの研究目的に支障が無ければある程度のコンタミは生じていても構わない。
そういう環境の中で今回の事件が発生している。小保方さんの研究は若山研のマウスで行われている。それ以前は彼女は自家繁殖マウスは使用していません。業者から買ったものだけです。
もし実験が購入マウスだけで行われていたら細胞の由来トレースはとても困難なものになっていたでしょうね。
若山さんがどういうマウス管理をしていたのかは分かりません。自家繁殖させていたらしいのは以下です。
①GOFマウス
②ロックフェラー大学で作ったB6-CAG-GFPホモマウス
③②からGFPを移し替えた129/Sv-X1-CAG-GFPホモマウス
④太田さんが岡部研から持ち込んだB6-Acr-CAG-GFPホモマウス
他のものは情報が無いのでわかりません。動物実験申請書では業者名が黒塗りされていて、恐らく自家繁殖分と書き分けられているので黒塗りされているのではないかと思われますが、詳細が分からない。
①は小保方さんが手記でばらしてしまった。②③は「僕のマウス」の嘘の言い訳をするために記者会見で自分でばらした。④も記者会見で岡部さんに譲ってもらって維持していたとばらした。手記では顕微授精もしていたような書きざまになっている。自家繁殖させているとお金の節約にはなります。
どの自家繁殖も業者のように兄妹交配を厳密にやっていればホモ率はどんどん上がってくる。でも自家繁殖でしかも研究目的からしてそれほど関係のないものなら、一つのコロニーで維持繁殖させる。すると近交系マウスから所謂クローズドコロニーマウスになる。親との戻し交配も発生しますから近交系マウスの原理通りにはホモ率は上がりません。それでも買った時の近交系は維持されています。
ここに違うマウスのコンタミが入るとどうなるかということです。白毛の129のコロニーの中に黒毛のB6が飛び込んで気づいて取り除いた時には既に交配していたとする。その子は全部混血色ですから生まれた時に取り除く。これでコンタミは取り除かれた。でも、いつ気づくかによりますね。生まれた子供が更に交配するほど長く気づいていないと、F2では白毛の子もいくつか残る。すると毛色以外の遺伝子がB6である白毛マウスが残ってしまう。マウスが成熟するまで1か月以上はかかりますから、こういうことはとても起きにくい。
今回のコンタミで顕著なのは③のマウスですね。B6から129へGFPだけを移し替えた。その手法は基本的にF1に対する129の戻し交配です。半分B6が入ったものでGFPの光っている子供だけに129を何度も交配させる。これも20継代続けると近交系になります。若山129近交系GFPマウスです。このマウスに調査時点でB6のSNPsが所謂近交系マウスとしては大量に入っていた。入っていたのは事実で、しかし、この原因が分からないわけです。
①戻し交配回数不足
②そもそものジャクソン研究所でのコンタミ残滓
③若山129近交系GFPマウス樹立後の新たなB6の飛び込み事故
もう一度確認しましょうかね。BCA報告の図です。
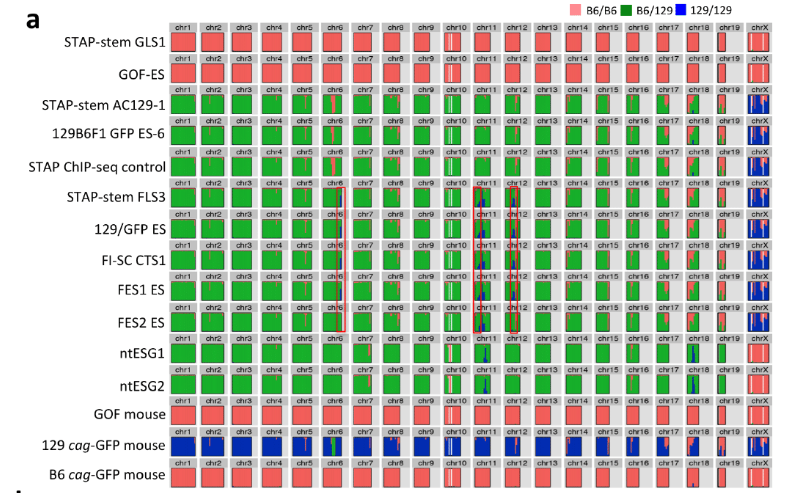
まず、ntESG1、G2の赤を見てください。ほとんどありませんが僅かに7,10,16番にあって他の細胞とも共有されている。これが②のジャクソン研究所でのコンタミ残滓ではないかと推測される。この129はterでテラトーマ専用に開発された129の亜種です。
それに対して、129cag-GFP mouseは129のX1です。市販のX1はデータとして示されていません。若山さんの「僕のマウス」の片割れの親マウスのデータだけが示されている。このコンタミはひどいですよね。市販されたものにこれだけコンタミがあるということはあり得ませんので、このコンタミは若山さんが作ってしまったものです。
一番特徴的な17,18,19番染色体を見ると上から順にAC129からFES2まで8ライン全部に共通していますね。
上3つは「僕のマウス」ESで捏造したと言ってるわけですから、一致しているのは分かりますよね。でも、更にFLSから下三つは太田ESで捏造したと言ってるんでしょ。更に下2つの太田ESはアクロシン入りの岡部マウスとのF1ですけど相手の129は市販のGFPのない129じゃなかったですか。というのも彼は最初自分はterと交配したつもりだったと証言していて、加えて、自分はB6に関してはアクロシン入りの岡部マウスしか使わないのでただ単にGFPと書いたと日経サイエンスに証言している。129側にもCAG-GFPの入った「僕のマウス」の片割れ129を使ったのならGFPに関してそれと注記するでしょう。
FES1、2のB6コンタミが他のラインの129側のコンタミと一致しているのは若山さんがFES1,2にFLS3を入れ替えたからです。私がなぜアルイミオウジ氏の最近の動向を報告したのか。このことがあるからです。彼の報告にGFPの有無検証が出るのを待ちましょう。
疑義は以下の解析は間違っていのだが、正しい答えは示されていないということです。

放医研は何を探していたのでしたかね。
CAG-GFPがどこに有るかですよね。放医研は最初15番にヘテロで入っていたと報告したことになっている。
でもそこにはGFPは無かった。実際には3番染色体にあったのです。岡部マウスのAcr-CAG-GFPは3番に有るんです。
ではどうして放医研は15番染色体にあると言ったのか。3番に有るのは内在性アクロシンプロモーター配列です。
では彼らは若山さんからプロモーター配列を知らされていてアクロシンプロモーター配列を含むプライマーで探していたのだ。若山さんは放医研が岡部マウスのGFPを見つけてくれると期待していたのに放医研は内在性アクロシンのプロモーターを捕まえてしまったのです。
ということはここにある18番にGFPが無いという結論も間違っている可能性がありますね。放医研はCAG-GFPを探しているのではなくてアクロシンGFPを探している結果になっている。18番にアクロシンプロモーターが無いのは当たり前です。ではCAG-GFPは調べ直したか。彼らは3番にAcr-CAGがあったと訂正した。では18番にヘテロで1コピーCAGが入っているか否かを調べたか。
少なくとも報告書は沈黙している。我々のSNPs解析の分析では129は「僕のマウス」の片割れであるという結果になっている。つまり18番にヘテロで1コピーのCAGがあるはずだと。
上の記者会見の説明映像のFLSはアクロシンプロモーターを探した結果です。右のAC129等はCAGプロモーターを探している。プライマーが違うんです。もしCAGプロモーターでFLSを調べていたら15番なんかに見つかるわけがない。そこにはCAGはありません。犯人は若山さんですよね。
(学さんの誤解4)今度は、
「すり替え、入れ替えなど、個人的な犯罪的行為をできるだけ考えないで、誰でも納得のいく説明はないものか?」という考察のようです。そこは最初に考えるところでないといけませんよね。まだだったんでしょうか。
互いに主張している事実関係に関して矛盾があるとどちらかが嘘をついていることになるんです。
ここをまず調べてないというところが怠慢ですね。
若山さんは「僕のマウス」を渡したと言っている。小保方さんは渡されたマウスでSTAP細胞を作って返したと言ってる。出来ている幹細胞を調べたらアクロシン入りだった。
若山さんが嘘をついているか、小保方さんが嘘をついているか、第三者が何かしたかのいずれかです。
この構図に関して一度も考えたことが無かったのでしょうか。
最初のキメラに第三者がESを入れて成功させてやるなんてことがあるでしょうか。いろんな種類の背景マウスですべて若山さんが渡したと主張しているマウス背景でない検査結果が出ている。第三者があり得ない決定的な証拠は最初の捏造が成功している捏造だからです。どうして成功していない段階から成功させるような手伝い捏造コンタミをする第三者がいますかね。この第三者は始めに成功させるようにコンタミさせた。ということはそれ以降も目的は成功させるようにコンタミさせていたのだということになる。どこにそんな第三者がいますか。
犯人は若山さんか小保方さんしかいないんです。そのことをまず押さえないと。選択肢が無くなったところから、ではどちらなのだと考える。
若山さんが嘘をついているのなら、彼の言ってることは一つも信じられない。小保方さんが嘘をついているのなら彼女の言ってることは皆嘘だ。二つを比較したらいいんですね。
彼女は手記でたくさんの証言をした。再現実験にも参加した。そして言ってることには嘘が無い。対して、若山さんは実験にも参加せず、矛盾に対する説明が無い。そして既存ESコンタミだとしている桂報告書がすべてだと証言した。つまり、既存ESがどこから来たのかということを証明しないと彼の証言は一つも信じられない。パートナー氏の質問状に彼は一切返事を書いてない。太田氏もそうです。どちらが嘘をついているかは明らかです。
では、どうしてこんなことになったか、という思考順序になっていくんですね。今頃「すり替え、入れ替えなど、個人的な犯罪的行為をできるだけ考えないで、誰でも納得のいく説明はないものか?」と考えているんですか?
(アルイミオウジ氏の最近の動向2)①SNPとSNVの違いをどう理解しているのか説明してもらいたい。
②129の特異的SNPとして登録されている箇所の数と、B6の特異的SNPとして登録されている箇所の数を教えて貰いたい。
③Ts.Marker 氏はそれぞれ1000万か所程度だが、理研が調べた箇所は129とB6で異なっている300万か所と言ってるが間違いないか。
>>
------ 追記 6/30 -----
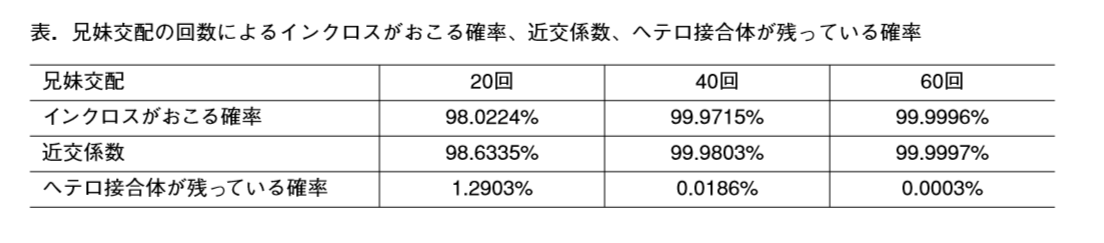
マウスのSNPsは1000万といわれていますが、調査報告ではB6と129で違う300万を使用している。④近交系マウスの定義は最低でもホモ率98%で、ヘテロ残存率1.3%ですから、30億dpに対して4000万弱のヘテロアレルがある。②はこの4000万アレルの中にはないはずだ。(あったらSNPs判定はできない。)アルイミオウジ氏の言うヘテロSNPsとは何を意味しているのか、答えてもらいたい。
>>
*www.anim.med.kyoto-u.ac.jp/Kuramoto/contents/ExpAnimGenet_1_inbred.pdf
⑤公開データ登録されている幹細胞のF1背景分に関して、129/Sv-X1にCAGがあるかないかのアルイミオウジ氏解析結果を教えて貰いたい。
(学さんの誤解5)
さて、ここまで説明後に初めて本来の話に戻れる。学さんの主張です。
>>
近交系は、クローンマウスではないので、98%で一致しても、残りの2%で一致しなければ、減数分裂で配偶子に多様性が生まれます。今300万箇所のSNPsの話をしている。ホモになっている場所だけの300万か所です。アルイミオウジ氏の話でも書きましたが、ヘテロのSNPを調べたらこのSNPs解析自体が意味をなさないでしょうよ。父母どちらもホモにそろっているからF1を造った時すべてヘテロになる。色分けだと緑になる。これが判定基準です。緑でなく青だとかピンクになっている部分があったら、それは親のSNPがヘテロだったということです。つまり、ホモであるべき場所がヘテロになっているから若山さんの維持繁殖させているマウスコロニーの近交系が崩れているということが分かる。本来ホモだという前提です。原理的にヘテロもあるのだとしたら、つまり登録されている特異的SNPがヘテロであり得るのだとしたらそもそもこの解析は何を識別していることになるでしょうか。根本的な無理解です。
自分の維持しているマウスコロニーにコンタミが無いかどうかを調べてくれる商売があるんですよ。登録されているデータがヘテロもあり得るんだったら調べようがないでしょ。登録されている特異的SNPsデータは全部ホモだというのが私の理解です。コンタミがあるとその場所がヘテロになる。
緑であるべきところがピンクになっているのは129にB6がコンタミしている。緑であるべきところが青になっているのはB6に129がコンタミしている。つまり、ピンクは129のコンタミを、ブルーはB6のコンタミを意味しているんです。区別が必要です。
129には以下の可能性がある。
a.市販の129/Sv-X1 GFP無し
b.市販の129/Sv-ter GFP無し
c.若山さんの自家繁殖させている129/Sv-X1 GFP無し
d.若山さんの自家繁殖させている129/Sv-ter GFP無し
e.若山さんの自家繁殖させている129/Sv-X1 CAG-GFP(ホモ)
B6には以下の可能性がある。
a.市販のB6 GFP無し
b.市販の(理研の)GOFマウス
c.若山さんの自家繁殖させているB6 GFP無し
d.若山さんの自家繁殖させているGOFマウス
e.若山さんの自家繁殖させているB6 CAG-GFP(ホモ)
f.大阪大岡部氏より移譲のB6-Acr-CAG-GFP(ホモ)
g.若山さんの自家繁殖させている岡部氏より移譲のB6-Acr-CAG-GFP(ホモ)
ピンクはa,c,dの可能性があるが、a.ではないはず。市販のものにこんなにコンタミの有ることは考えにくい。ntESG1,G2はb.dのどちらかだが、ほとんどコンタミは無い。
ブルーは全部gです。
もう一度貼り付けましょう。
ピンクはすべてに共通しているが6番染色体のみが「僕のマウス」グループと違っている。ここは129 cag-GFP mouseは緑になっている部分です。
それに対してブルーは元の岡部マウスのデータが無いが、「僕のマウス」グループ以外は全部自家繁殖の岡部マウスです。ここにコンタミのある部分が青で出ている。その出方に3種類ある。
①FLS3,129/GFP ES.CTS1,FES1
②FES2
③ntESG1、G2
なぜ3種類になるのか。まず最初に作られたのは③です。太田さんが岡部研から持ってきたときの岡部マウスで作ったもの。11番と18番にコンタミがある。
②は11番に③とは別のコンタミがある。ただし、この129はterではありません。なぜなら129 cag-GFP mouseのピンクパターンが共通しているからです。
①は6,11,12番に共通のコンタミが見られる。しかも11番は②と共通している。
以下が、学さんが言いたいことですね。我々の長年の関心事でもある。
>>
もっとも、調査チームが、FES1 FES2問題を論じているのは、ここではないけどね。
調査チームは、幹細胞成立後に起きた塩基変異が焦点だ。
何度も出てくるが、桂報告書の以下の6頁部分が、FES1 FES2問題の鍵となると思う。今回 4 種の幹細胞には、第 6、第 11、第 12 染色体上に 129 に特徴的なクラスターが、また第 17、第 18、第 19 染色体等に C57BL/6 のクラスターが 認められることから、TaqMan PCR によって観察された 129 ホモの SNPs はこれら幹細胞 の作製に使用したマウスに存在した遺伝的背景の不均一性によるものと結論づけた。 ES 細胞 FES1 と FES2 でのみ異なる SNPs に関して、両者の遺伝的背景の相違によると 判断された上記第 6、第 11、第 12 染色体の SNPs クラスターを除外し、残った 1,290SNPs を用いて比較を行うと、STAP 幹細胞 FLS3、FI 幹細胞 CTS1、および、ES 細胞 129/GFP ES は同一細胞株といって良い程の高い類似性を示すことが判明した。従って、STAP 幹細胞 FLS3、FI 幹細胞 CTS1、および 129/GFP ES は同一の細胞由来であり、ES 細胞 FES1 と同 一、あるいはそれから派生した株の可能性が高い、と結論づけた。 FES1 FES2の作製時、親マウスがどのような状態であったかはわからない。
但し、同じ受精卵由来でないことはわかるが、両者のSNPが離れている原因が謎になっている。学さんの疑義は今述べた我々の疑義そのものですね。
そして学さんはその原因を以下のように考えるのだけれども、日本語の助詞の使い方が間違っていてとても誤解をうけやすいのと、もう一つは推測そのものに間違いがあってややこしい。韓国系の人は助詞の間違いは少ない。シナ系の人は助詞の使い方が難しくてよく間違えますね。
>>
それぞれ、129もB6の両マウスは、マウス交雑の結果ヘテロであることから、F1において、129/B6とならずに、129/129、B6/B6の組み合わせができうる。
2003年に若山研究室に入導入されたアクロシン入りB6マウスは、FES1 FES2の親となったが、その遺伝子の詳細は公表されておらず、やはり129マウスのコンタミは起きていた。
つまり、FES1 FES2のSNPの様相は、ヘテロな親から引き継いだものと、ES作成後に生じてきた1塩基変異を合わせて考える必要がある。学さんの今までの考えを理解すると、上の文章は以下のように直せる。
>>
それぞれ、129も
(→と)B6の両マウスは、マウス交雑
(→混入)の結果
(特異的SNPsが一部)ヘテロであることから、F1において、129/B6とならずに、129/129、B6/B6の組み合わせができうる。
2003年に若山研究室に入導入されたアクロシン入りB6マウスは、FES1 FES2の親となったが、その遺伝子の詳細は公表されておらず、やはり129マウスのコンタミは起きていた。
つまり、FES1 FES2のSNPの
(異なった)様相は、ヘテロな親から引き継いだものと、ES作成後に生じてきた1塩基変異を合わせて考える必要がある。
ここでも「も」の使い方がおかしいですね。学さんはシナ系の一世のとてもよく日本語のできる方ということでしょうかね。帰化人の2世で日本の学校に通った子はこういう間違いは絶対にしません。周りの子供たちから矯正されてしまいます。日本人がこのような助詞の使い方の間違いを絶対にしないのも同じ理由です。
でも、外国人、もしくは帰化1世の人だと分かれば簡単に理解できます。我々は日本語はプロですからね。事情が分かれば推測が可能になる。彼女が言ってることは我々の言ってることと全く同じです。
問題は彼女の推測自体が正しいかということです。「ES作成後に生じてきた1塩基変異を合わせて考える必要がある。」ということですが、上の図を見て、そんな推測ができますか。ntESG1G2にあった青はどうして他の細胞では消えてしまっているのですか。これは元のマウスが違うとしか思えませんね。というよりも、この実験が最初ですから、他の岡部マウスからこの時の青が消えた理由が解明されないといけない。加わったものより消えた方が問題でしょう。
岡部マウスは太田さんが若山ラボに来たことが契機で大阪大から移譲された。太田さんはその岡部B6マウスを使って、ntESG1,G2とFES1,2を作っている。
2005/1/20に凍結し、2007年にもう一度解凍して使用し再度2007/8/3に凍結したntESG1,G2にあったブルー部分のヘテロSNPsが、2005/12/7に凍結したFES1,2からは消えている。(Ooboeさん情報)途中ですが、今の考察と関連しますから、学さんのところに書き込まれたOoboeさんからの情報をここで確認しておきましょう。「Ooboeさんとパートナー氏の資料館」にもアップしています。
>>
現在、FES1、とFES2の調査用のES細胞サンプルの考察が続いておられますが、パートナーが入手しているそのサンプルについての情報をお伝え致します。参考にして下さいませ。
2019/12/04 URL 編集
Ooboe
前コメント、名前入力わすれました。ごめんなさい。
FES1などについてのパートナー入手情報
理研広報は、調査サンプルを受領し、解析を担当した、CDB非対称細胞分裂研の研究者に直々に確認され、その研究者はFES1とntESG1は山梨大、若山教授から取り寄せました。
FES2とntESG2は山梨、若山教授とは別のところから取り寄せました。
これと同じ内容は、情報公開室が確認しています。
今晩、続きをお伝え致します。
2019/12/04 URL 編集
Ooboe
続きです。
前記にありますように、理研広報や調査委員会事務局に調査用サンプルの取り寄せについて報告された、BCA論文の筆頭著者でもあった研究者は、BCA論文中では、大田氏からFES1、FES2、ntESG1、ntESG2、を取り寄せたことになる記述をしています。
世界に発信されたBCA論文と、食い違う内容報告を、理研広報など、理研組織内では共有確認しているわけですが、なにより取り寄せ依頼し、受領し、解析したご当人がこの食い違いについての諸々の事実を把握していると思われます。
2019/12/04 URL 編集
Ooboe
さて、調査用ESサンプルを取り寄せ依頼、受領、解析をされた、CDB研究者はなにゆえに、理研にとっての公式発表を担当する広報室や、法令遵守担当のコンプライアンス室などの対し、組織内部にだけに共有される情報を伝え、対外発表のBCA論文や、調査報告書とは食い違う報告をしたのでしょうか?
もし、広報などに伝えた、内容通りならFES1とntESG1は、山梨大で用意された調査用サンプルということになります。
そして、FES2と、ntESG2は、別のところということですから。FES1とFES2は、出所が違う訳であり、細胞作成された、場所も同じでない別々に作られた、ことになります。又は、株分けでしょうか?
いずれにしろ、FES1は、京都大、大田氏から、取り寄せたのではなく、山梨大、若山研から取り寄せたと、BCA筆頭著者は、理研内部組織に伝えていたこの事実は、客観的公的事実であることには、変りがございません。
2019/12/05 URL 編集
Ooboe
パートナーは、FES1などのサンプル受領についての、MTA契約文書の開示請求をしましたが、情報公開室は、解析担当研に確認した結果を、パートナー宛に回答書を送付しています。その記述の中に、解析担当研究者が若山教授に直接依頼したもので、MTAや取り寄せ手続きの公的文書等は交わしていません、とありました。
6月30日理研は機関として、コンプライアンス室が予備調査を開始するとの発表をしていますが、しかしFES1等の調査取り寄せに予備調査事務局は、関与していませんでした。
この事案は普通の研究者間でのサンプル提供、受領手続きではありません。機関として、公的に取り寄せるべき事案であった訳ですから、関与していれば公的事務局として、文書交付は為されたはずでした。
解析調査をする側は、サンプルを取り寄せの依頼するのですから、取り寄せ宅配経費は、依頼側が支払うはずです。しかし解析担当研は会計処理をされてませんし、又、山梨大学、若山研の会計処理も為されてませんので、若山氏が自費でFES1の宅配経費負担をして理研解析担当研究者に送付していたことになります。
ということから、依頼したのは、逆に若山氏側であった。というのが整合的に帰結されてくる訳です。
以上
FES1等については不明瞭な事実が更に重なって存在していますが、省きます。
いずれにいたしましても、真正な調査サンプルとしての資格が無いに等しいのが、 FES1等サンプルでありました。
2019/12/05 URL 編集ほぼすべて既知情報ですが、新しい情報の一つは直接やり取りした人がDaijiro Konnoさんだったということですね。でも指示は責任著者の松崎氏から出ますからね。
「BCA論文の筆頭著者でもあった研究者は、BCA論文中では、大田氏からFES1、FES2、ntESG1、ntESG2、を取り寄せたことになる記述をしています。」の部分、事実であるかどうかは別として、そもそも太田氏が出発点だということになっているから、訴訟法的な厳密な確認はしないで、そう書いたという言い訳はできます。変だと分かっていたろうがと責めることはできますけどね。
「FES1等の調査取り寄せに予備調査事務局は、関与していませんでした。」というのは重大です。ガヴァナンスが無いんですね。
ところで、今ため息ブログ氏がこの件に関して以下のように書いている。
>>
近交系に混入があってもはや近交系ではなくなり遺伝背景が均一でない、これでFES1とFES2のSNPsの違いを説明できると言っているんでしょ。それだけでは説明できないと学さんも言っていて、我々も今、ここでそれを考えているところですね。ため息氏は全然専門家でないし、また、素人であってもこの件に関してよく調べている人でもないんで、こんな人と議論していても意味無いでしょうと言ってるんですけどね。
ため息氏のブログを読んでいたら目にとまったのでコメントしておきましょうかね。同じように誤解している人は多そうです。
>>
今さら何を言っているのでしょうね。FLSもAC129と同じようにCAG-GFP遺伝子の挿入部位を読んだのです。しかし、アクロシンプロモーターが付いたGFP遺伝子とCAG-GFP遺伝子が連結していたため、本来読むべき配列ではなく、通常15番染色体にあるアクロシンプロモーターの配列を読んでしまい、「15番」と誤認したのです。15番にはCAGは無いと言ってるのに分からないんですね。アクロシンプロモーターを含むプライマーで探したから15番の内在性アクロシンプロモーターを誤認した。CAGのプロモーターを探していたら15番にはCAGはないから誤認するわけないでしょ。
この人はもと数字君とコンビの岡目何とか君じゃなかったかな。
何度でも貼り付けましょう。
右はCAGプロモーターを含むプライマーで挟んだ。
左はアクロシンプロモーターとCAGプロモーターの両方を含むプライマーで挟んだ。
左のサンプルを右のプライマーで挟んだら15番にはCAGプロモーターは無いから検出できない筈だよと言ってる。アクロシンプロモーターとCAGプロモーターの両方を含むプライマーで挟んだ時にのみ誤認があり得る。しかし、この場合左の18番に無いとされている結果もアクロシンプロモーターが無いだけという可能性があると言ってる。
そもそも、右はCAGプロモーターしか含んでいないプライマーなのは15番に誤認がないのでわかる。ではどうして左はアクロシンプロモーターを含むプライマーで探したのかね。分かってたんでしょ。アクロシンを3番に発見してもらいたかったけど15番と間違えたので遠藤様が登場したということでしょ。
放医研のお友達にはどういうゲノムウォークをしたから間違えたのだとちゃんとした説明をして欲しいね。無料だから間違えたのかね。この人はタカラバイオでは商売にならないから雇われないだろうな。こんな間違いしてたら、損害賠償請求されてしまう。小保方さんの弁護士は何をしてたのかね。それとも放医研の人は無関係だったのかな。
(学さんの誤解6)ここ2回の記事で学さんは立ち直ったようです。もう彼らの相手はせずに、問題そのものを相手にするということのようです。直近の最後の記事に関しては私は完全同意です。そういう姿勢であっちを相手にしないと決めてくれたら私はそこに戻って質問してもいいですね。私は学さんの知識に関しては疑ってない。特にTCRについて教えられたことと、丹羽さんのGFPはクレだと私の勘違いを注意してくれた。忘れてませんよ。
時々刺激して無いと寝込んじゃうという意味では他の人がやってくれますよ。私なんか彼らが工作を続けていてくれているから何度でも繰り返し、若山さんが犯人だと繰り返せるわけです。彼らが続ける限り永遠に若山さんが犯人だと書き続ける。どうしてかというと彼らがいつまでも嘘の工作をし続けているからです。彼らが永遠に黙り込んじゃったらどうしますか。誰も何も言ってないのに、一人で若山さんが犯人だと同じ事を何度も繰り返さなくてはならなくなる。そうなると読者はそれはもう聞いたよと読まれなくなってしまう。雑談コーナーは今黙ってしまった。気づかれたか。はは。
(学さんが相手しなくていいように)>>
一ヶ月前の質問です。まだ生きています。答えて頂戴。
1) 学とみ子説「T細胞はキメラに寄与できない」のだから、TCR再構成を使う実験の提案者(西川氏)やそれを受けた笹井氏等は免疫学・発生学・細胞学に無知な方だったの?
2) FES1、FES2についてとは何を問題にしている?
3) 桂報告書のどこにES説では成り立たないことが書いてあるの?
4) 「科学的事実は、その時から、全く変化しました。」→ 5年経過してどんな科学的事実が変化したの?
5) 「これだけの事実が出てきて」→ 5年経過してどんな事実が出てきたの?
6) 「桂報告書に、若山研究室の見取り図を載せた人と同じレベルです。」→ どんなレベル?
7) 「遺伝子状態」て何?どのような状態のこと?
8) 「遺伝子が欠失すると多能性が発揮される」→ 具体的なそのような例を示して頂戴。核移植とiPSは遺伝子欠失ではないよ、そして実験的な多能性の発揮はこの2つしかないのでは?
9) 女王様とフィンガーボールのエピソードは笑い話だったの?
10) 幹細胞作成時には細胞をソートしていないと思うのですが、ソートしているとはどういうことでしょ?1) 学とみ子説「T細胞はキメラに寄与できない」のだから、TCR再構成を使う実験の提案者(西川氏)やそれを受けた笹井氏等は免疫学・発生学・細胞学に無知な方だったの?
学さんは論文どうりのSTAP細胞(小保方細胞ではない)があるという立場で考えておられるので、その推測の中でT細胞は選別されて生き残ってない筈だと考えたんです。白血球の他の細胞が生き残ったのではないかと。
2) FES1、FES2についてとは何を問題にしている?
私も今問題にしていますが、私の場合は細胞が入れ替えられているという基本的視点からの問題意識です。言うまでもありませんが細胞の入れ替えが無いのなら犯人は小保方さんになります。特にGLSについては動きませんね。ntESG1G2は論文、ラベルと中身が違っている。細胞の入れ替えがあったことを示唆している。Ooboeさんたちの追っている問題です。学さんは若山さんが犯人だということを考えていないので、なかなかすべての情報要素を整合的に整理できないでいる段階だとみています。
3) 桂報告書のどこにES説では成り立たないことが書いてあるの?
そんなことは書いてない。既存ESだと言ってます。ただ、キメラの捏造を断定していない。それは私に言わせると政治的配慮なんですが、学さんは調査チーム内、或は理研内での対立を想像している、特に竹市さんは15番にGFPのあるマウスを調査中と報告している。理研には小保方さんを雇って問題を拡大した責任に関して各所に配慮があるという事情もある。そもそも大した問題でなかったものです。
4) 「科学的事実は、その時から、全く変化しました。」→ 5年経過してどんな科学的事実が変化したの?
5) 「これだけの事実が出てきて」→ 5年経過してどんな事実が出てきたの?
桂報告書記者会見の後に、より詳しいBCA報告がでて、ワトソン氏はそれを知らずに私と議論して、最後にブレンデルのピアノに関して喧嘩別れしましたが、あれはTs.Marker氏によるとDORAさんだったらしい。はは。その後再現検証実験についての丹羽さんの報告と相沢さんの報告がでた。米国でも再現実験の報告が出た。7月頃にはキンガ・ヴォイニーツの論文がでてティシュー論文が批判的に引用された。それといろんな人が既存の科学的事実から演繹した知見も増えてきているというような意味でしょう。
6) 「桂報告書に、若山研究室の見取り図を載せた人と同じレベルです。」→ どんなレベル?
アホでしょ。何も意味してない。
7) 「遺伝子状態」て何?どのような状態のこと?
これはこの言葉の使われた状態を知らないので分からない。
8) 「遺伝子が欠失すると多能性が発揮される」→ 具体的なそのような例を示して頂戴。核移植とiPSは遺伝子欠失ではないよ、そして実験的な多能性の発揮はこの2つしかないのでは?
2つ以外にミューズ細胞とムーさんの細胞があるかな。小保方細胞が何なのかは研究中ですね。今報告は止まってるが。ただ、「遺伝子が欠失すると多能性が発揮される」というのはどういう文脈の中で使われたのか、多能性獲得というのはリプログラムされることなので、欠失すると遺伝子情報が不完全になるという意味なので、遺伝子欠失と多能性発揮を結び付けた研究は誰もまだしてないのではないか。なにしろ、学さんはSTAP細胞有りの前提で考え続けられている過程での試行錯誤ではないでしょうかね。
9) 女王様とフィンガーボールのエピソードは笑い話だったの?
暗喩というのは何を裏に隠すかということなので、意図は本人でないと分からないし、分からないときに無理にわかる必要もない。何事かをあからさまにしたいときには比喩は使わない。
10) 幹細胞作成時には細胞をソートしていないと思うのですが、ソートしているとはどういうことでしょ?
以前から気になってましたが、木星リストのFLBはソートされていると書かれているんです。まず事実を確認して後に、このソートは何かということになる。我々は無論ntESをキメラ胚に入れた後にそれを幹細胞としたわけですが、ここにはリシピエントのインナーセルマスが残るので、GFPでそれをソートしたと考えている。テラトーマの体細胞はソート前の幹細胞を若山さんが上から注射したからリシピエントのインナーセルマス由来のES細胞がテラトーマ化したとみている。これは2Nでも4Nでもソートされている。4Nでも胎生致死以前の段階なのでまだ死んでないと見られている。和モガさんも考察しているところです。
- 2019/11/25(月) 09:40:06|
- AC129
-
-
| コメント:11












