theartofintelligence.blog.jp/archives/5447318.html
新設しました。
便所の落書きではありません。ここはここでヒソヒソ話の場です。ひひひ。
- 2020/01/30(木) 07:48:04|
- 新設・STAP事件簿
-
-
| コメント:8
venonasecrets.blog.jp/archives/5427059.html
古代史も引っ越ししました。
2020/2/7に過去記事の引っ越し終了しました。
これからは新記事です。
- 2020/01/28(火) 15:12:22|
- 引っ越し・古代史ブログ
-
-
| コメント:0
(英文)
Expression analysis by chromosome to identify trisomy
FPKM values originating from ESCs (SRR1171574 and SRR1171575) and STAP cells (SRR1171578 and SRR1171579) were calculated, and genes having >0.01 FPKM in all four original experiments were selected. Genes without pseudogenes were classified for chromosomes, and the log ratio of the average of two experiments using the same cells was determined for each chromosome. The distribution of relative FPKM values was evaluated using one-sided t-test against the mean log ratio of whole genes.
トリソミー識別のための染色体による発現解析
ES細胞(SRR1171574とSRR1171575)及びSTAP細胞(SRR1171578とSRR1171579)由来のFPKM値が算出され、全4回のオリジナルの実験における0.01以上のFPKMを有する遺伝子が選択された。染色体上に偽遺伝子を伴わない遺伝子を分類し、各染色体について同じ細胞を用いた2つの実験の平均の対数比を決定した。相対FPKM値の分布は、全体の遺伝子の平均log比に対して、一群t検定を用いて評価した。
(英文)
MEF marker genes
Marker genes of feeder cells were identified using unpublished RNA-seq data provided by Dr Jafar Sharif and Dr Kyoichi Isono. Gene expression differences among ESCs, TSCs and MEFs were compared using the cuffdiff program, and genes that were significantly highly expressed in MEFs were selected. Genes encoding cytokines and extracellular matrix-related genes were selected to illustrate the features of feeder cells in Fig. 2E.
MEFマーカー遺伝子
フィーダー細胞のマーカー遺伝子はジャファルシャリフ博士や磯野恭一博士によって提供されている未公開のRNA-seqデータを用いて同定されている。 ES細胞、TS細胞及びMEF(マウス胎児線維芽細胞)間の遺伝子発現の違いはcuffdiffプログラムを用いて比較されていて、MEFの中で有意に高頻度で発現された遺伝子が選択されている。サイトカインおよび細胞外マトリックス関連遺伝子をコードする遺伝子が図2Eのフィーダー細胞の特徴を説明するために選ばれている。
(英文)
Acknowledgements
I would like to acknowledge Dr Akihiko Yoshimura, Keio University, who initially suggested on his website that NGS data could be evaluated by SNP analysis. Dr Ichiro Taniuchi and Dr Nyambayar Dashtsoodol, RIKEN-IMS, offered insightful information for interpreting the experimental procedures. Dr Norihito Hayatsu, RIKEN-IMS, provided unpublished inbred/outbred mice sequences to validate the genotype analysis. Dr Jafar Sharif and Dr Kyoichi Isono, RIKEN-IMS, provided unpublished transcriptome data to identify marker genes. Dr Shinichi Nakagawa, RIKEN, provided critical comments on my manuscript, and Mr David Gifford, RIKEN-CSRS, edited the text. I especially thank the authors of the retracted paper, Dr Teruhiko Wakayama, University of Yamanashi and Dr Hitoshi Niwa, RIKEN-CDB, for commenting on this manuscript.
謝辞
私はまず慶應義塾大学の吉村明彦博士に謝辞をささげたい。博士は最初にウェブサイト上でNGSデータがSNP解析により評価しうると示唆された方です。理化学研究所統合生命医科学研究センターの谷内一郎博士とNyambayar Dashtsoodol博士には実験手順を解釈するための洞察力に富んだ情報を提供していただきました。同じく理化学研究所統合生命医科学研究センターの早津徳人博士には遺伝子型解析を検証するために未発表の近親交配系/非近親交配系のマウスシーケンスを提供していただきました。同じく理化学研究所統合生命医科学研究センターのジャファルシャリフ博士と磯野恭一博士にはマーカー遺伝子を特定するために未発表のトランスクリプトームデータを提供していただきました。理研の中川真一博士には私の原稿に重要なコメントを提供していただきました。そして環境資源科学研究センター (CSRS)のデビッド·ギフォード氏はテキストを編集してくれました。また特に取下げ論文の著者である山梨大学の若山照彦博士と理研CDBの丹羽仁博士にはこの原稿について論評していただいたことに感謝致します。
(英文)
References
Ben-David, U., Mayshar, Y. & Benvenisty, N. (2013) Virtual karyotyping of pluripotent stem cells on the basis of their global gene expression profiles. Nat. Protoc. 8, 989–997.
Chang, G., Gao, S., Hou, X. et al. (2014) High-throughput sequencing reveals the disruption of methylation of imprinted gene in induced pluripotent stem cells. Cell Res. 24, 293–306.
DeVeale, B., van der Kooy, D. & Babak, T. (2012) Critical evaluation of imprinted gene expression by RNA-Seq: a new perspective. PLoS Genet. 8, e1002600.
Gropp, A. (1982) Value of an animal model for trisomy. Virchows Arch. A Pathol. Anat. Histol. 395, 117–131.
参照
Ben-David, U., Mayshar, Y. & Benvenisty, N. (2013) 『世界的遺伝子発現プロファイルに基づく多能性幹細胞の仮想核型分類』 Nat. Protoc. 8, 989–997
Chang, G., Gao, S., Hou, X. et al. (2014) 『大量処理シークエンシングが誘導多能性幹細胞の中のインプリンティング遺伝子のメチル化破壊を明らかにする』 Cell Res. 24, 293–306
DeVeale, B., van der Kooy, D. & Babak, T. (2012) 『RNA-配列によるインプリント遺伝子発現の重要な評価:新たな視点』PLoS Genet. 8, e1002600
Gropp, A. (1982) 『トリソミーのための動物モデル』 Virchows Arch. A Pathol. Anat. Histol. 395, 117–131
(英文)
Hussein, S.M., Batada, N.N., Vuoristo, S. et al. (2011) Copy number variation and selection during reprogramming to pluripotency. Nature 471, 58–62.
Kim, Y.M., Lee, J., Xia, L., Mulvihill, J.J. & Li, S. (2013) Trisomy 8: a common finding in mouse embryonic stem (ES) cell lines. Mol. Cytogenet. 6, 3–7.
Lagarrigue, S., Martin, L., Hormozdiari, F., Roux, P.F., Pan, C., van Nas, A., Demeure, O., Cantor, R., Ghazalpour, A., Eskin, E. & Lusis, A.J. (2013) analysis of allele-specific expression in mouse liver by RNA-Seq: a comparison with cis-eQTL identified using genetic linkage. Genetics 195, 1157–1166.
Liu, X., Wu, H., Loring, J., Hormuzdi, S., Disteche, C.M., Bornstein, P. & Jaenisch, R. (1997) Trisomy eight in ES cells is a common potential problem in gene targeting and interferes with germ line transmission. Dev. Dyn. 209, 85–91.
Mayshar, Y., Ben-David, U., Lavon, N., Biancotti, J.C., Yakir, B., Clark, A.T., Plath, K., Lowry, W.E. & Benvenisty, N. (2010) Identification and classification of chromosomal aberrations in human induced pluripotent stem cells. Cell Stem Cell 7, 521–531.
Hussein, S.M., Batada, N.N., Vuoristo, S. et al. (2011) 『多能性再プログラミング中のコピー数の変化と選択』 Nature 471, 58–62
Kim, Y.M., Lee, J., Xia, L., Mulvihill, J.J. & Li, S. (2013) 『トリソミー8:マウス胚性幹(ES)細胞株における一般的所見』 Mol. Cytogenet. 6, 3–7
Lagarrigue, S., Martin, L., Hormozdiari, F., Roux, P.F., Pan, C., van Nas, A., Demeure, O., Cantor, R., Ghazalpour, A., Eskin, E. & Lusis, A.J. (2013) 『RNA-配列によるマウス肝臓における対立遺伝子特異的発現の分析:遺伝子連鎖を使って同定されたcis-eQTLとの比較』 Genetics 195, 1157–1166
Liu, X., Wu, H., Loring, J., Hormuzdi, S., Disteche, C.M., Bornstein, P. & Jaenisch, R. (1997) 『ES細胞におけるトリソミー8は遺伝子ターゲッティングにおける共通の潜在的な問題であり、生殖系列伝達を妨害する』 Dev. Dyn. 209, 85–91
Mayshar, Y., Ben-David, U., Lavon, N., Biancotti, J.C., Yakir, B., Clark, A.T., Plath, K., Lowry, W.E. & Benvenisty, N. (2010) 『人間の人工多能性幹細胞における染色体異常の同定及び分類』 Cell Stem Cell 7, 521–531
(英文)
Obokata, H., Sasai, Y., Niwa, H., Kadota, M., Andrabi, M., Takata, N., Tokoro, M., Terashita, Y., Yonemura, S., Vacanti, C.A. & Wakayama, T. (2014a) Bidirectional developmental potential in reprogrammed cells with acquired pluripotency. Nature 505, 676–680.
Obokata, H., Wakayama, T., Sasai, Y., Kojima, K., Vacanti, M.P., Niwa, H., Yamato, M. & Vacanti, C. (2014b) Stimulus-triggered fate conversion of somatic cells into pluripotency. Nature 505, 641–647.
Tanaka, S., Kunath, T., Hadjantonakis, A.K., Nagy, A. & Rossant, J. (1998) Promotion of trophoblast stem cell proliferation by FGF4. Science 282, 2072–2075.
Wang, L., Wang, S. & Li, W. (2012) RSeQC: quality control of RNA-seq experiments. Bioinformatics 28, 2184–2185.
Obokata, H., Sasai, Y., Niwa, H., Kadota, M., Andrabi, M., Takata, N., Tokoro, M., Terashita, Y., Yonemura, S., Vacanti, C.A. & Wakayama, T. (2014a) 『取得多能性を持つ再プログラム細胞における双方向への発生能力』 Nature 505, 676–680
Obokata, H., Wakayama, T., Sasai, Y., Kojima, K., Vacanti, M.P., Niwa, H., Yamato, M. & Vacanti, C. (2014b) 『体細胞の多能性への刺激惹起性運命変換』 Nature 505, 641–647.
Tanaka, S., Kunath, T., Hadjantonakis, A.K., Nagy, A. & Rossant, J. (1998) 『FGF4による栄養膜幹細胞増殖の促進』 Science 282, 2072–2075
Wang, L., Wang, S. & Li, W. (2012) 『RSeQC:RNA-seq実験の品質管理』 Bioinformatics 28, 2184–2185
(英文)
Supporting Information
(Filename) gtc12178-sup-0001-FigS1.pdf
(Format) application/PDF
(Size) 265K
(Description) Figure S1 Allele distributions from the RNA-seq data obtained for the cell lines reported in the Obokata et al. study. CD45+ cells (gray), ESCs (yellow), STAP cells (blue), STAP-SCs (green), TSCs (orange), EpiSCs (light blue), and FI-SCs (red). The ESCs, STAP cells, STAP-SCs, FI-SCs, and epiblast stem cells (EpiSCs) were annotated as being derived from a 129B6F1 strain, and the TSCs as from a CD1 strain.
サポート情報
(ファイル名) gtc12178-sup-0001-FigS1.pdf
(書式) application/PDF
(サイズ) 265K
(説明) 図S1 小保方らの研究の中で報告された細胞株に対して得られたRNA-seqデータからの対立遺伝子分布。 CD45+細胞(灰色)、ES細胞(黄)、STAP細胞(青)、STAP幹細胞(緑)、TS細胞(オレンジ)、EpiSCs<エピプラスト幹細胞>(水色)、およびFI幹細胞の(赤)。 ES細胞、STAP細胞、STAP幹細胞、FI幹細胞、および胚盤葉上層幹細胞(EpiSCs)は129B6F1株由来のものとして、またTS細胞はCD1株由来のものとして注釈されている。
(英文)
(Filename) gtc12178-sup-0002-FigS2.pdf
(Format) application/PDF
(Size) 214K
(Description) Figure S2 Allele frequencies of all chromosomes. SNPs on all autosomes and the X chromosome were counted, and their distributions are indicated. The RNA-seq data from the CD45+ and STAP cells are identical to those used in Fig. 3. Numbers after each chromosome name are those of applied SNPs of CD45+ rep1, CD45+ rep2, STAP rep1, and STAP rep2, respectively.
(ファイル名) gtc12178-sup-0002-FigS2.pdf
(書式) application/PDF
(サイズ) 214K
(説明) 図S2 すべての染色体の対立遺伝子頻度。すべての常染色体とX染色体上のSNPが計数され、かつそれらの分布が示されている。 CD45+およびSTAP細胞からのRNA-seqデータは図3で使用されているものと同一である。各染色体名の後の数字はそれぞれ、CD45+ rep1、CD45+ rep2、STAP rep1、及びSTAP rep2のSNPの数である。
(英文)
(Filename) gtc12178-sup-0003-TableS1.xlsx
(Format) application/msexcel
(Size) 11K
(Description) Table S1 RNA-seq raw data used in this study
Please note: Wiley Blackwell is not responsible for the content or functionality of any supporting information supplied by the authors. Any queries (other than missing content) should be directed to the corresponding author for the article.
(ファイル名) gtc12178-sup-0003-TableS1.xlsx
(書式) application/msexcel
(サイズ) 11K
(説明) 表S1 本研究で用いたRNA-seqの生データ
ご注意:ワイリーブラックウェルは、著者によって提供されるあらゆる補助情報の内容や機能についての責任を負いません。(コンテンツの欠落を除く)ご質問は記事の責任著者の方へお願いします。
- 2020/01/27(月) 11:39:31|
- 遠藤論文
-
-
| コメント:0
thomasmcknight.livedoor.blog/archives/5614292.html
ふむ。便所の落書きはSTAP事件だけでいい。ひひひ。
- 2020/01/26(日) 21:40:47|
- 引っ越し・釣りブログ
-
-
| コメント:0
さて、考察である。
取り敢えず、Ts.Marker 氏は自分の発見事実をブログで公開した。和モガ氏やアルイミオウジ氏とも連絡を取られている様子なので、裏ではある程度の意見交換があるのかもしれないが、一般の読者としては分からないから、ただ、事実報告しかない状態である。
それに対して、和モガ氏はこの事実に関する自分の解釈を書かれている。事件なのであるから、事件視点で何が行われたから、こういう科学的事実が出ているのかと問うことになる。
最初の推測はどこかの時点でクローンが作られているという推測から、ドナー核の周りにわずかに付着している細胞質からリシピエントの細胞質にミトコンドリアが混入するという推定をした。
ただし、和モガ氏のクローン説は日程検討に誤りが入っていて、私は既にそれを批判しているが修正はいまでにされていない。だからクローンがなぜ使われるのかの説明は疑義の入ったままに、その可能性を論じている事になるので、事件との絡みを離れている。
それを前提にして、それぞれの幹細胞はntESであり、インジェクション時にドナーの細胞質からわずかに入ったミトコンドリアがAの原因だとした。
>>
調査委員会はFLS3はFES1由来で、FES1は129×B6の受精卵ES細胞ということであった。それなら、mtDNAの12,188は②のように塩基Tで灰色表示されるはずである。作製者の太田氏は作成後、研究には使わなかったというので、突然変異が半分まで蓄積するはずもない。FLS3が受精卵ES細胞でないのは明らかだ。ここで、太田ESの嘘が暴かれた。太田ESは他にも嘘が多くて、証言に矛盾がある。
しかし、太田氏が使った129/Sv X1が市販のものであったのなら、調査現在のFES1の中身はその時の太田ESではないということになる。
ただ自家飼育の129/Sv X1のコロニーが維持されていたのなら、クローンであるとは言い切れない。言うまでもないが、小保方さんがFES1を渡したからFLSができたなんて話はもっと言いきれないことになる。要するに何にも調べられないまま、決めつけの結論を書いただけなのが桂報告書なのである。
そのことはさておいて、ここで和モガ氏は勘違いしている。リシピエントはICRである蓋然性が高い。今までの実験やその申請書でICRが書かれている。従って、市販のきれいなマウスだとTベースである。ここにAとして何が入ったのかという問題になる。和モガさんは逆を書いている。
>>
AとTが両方でるのは、FLS3が体細胞クローンで12,188番のTの塩基はドナーの129B6F1の129系塩基T(②)に、レシピエントのmtDNA(おそらくICRマウスで塩基はA)が混在しているからだと思われる。
しかも、加えてTs.Marker さんの調査ではICRはデータベースではTであった。
事実はTの中にAが入ったというだけのことである。まずは原因をクローン胚インジェクション操作だということに限定しないで考えた方が一般的考察になる。
和モガさんはSNPsを持ち出してきているが、核DNAでしかも近交系マウスに関して特に99%ホモになっているローカスのその近交系マウスに特異的SNPsが登録されている。そしてミトコンドリアDNAは母系一本しかないので、その意味のヘテロは無いので紛らわしい。ただし、そのマウスのDNA配列は全部読まれていて、あの場所がAであるマウスはTs.Markerさんが調べた限りではNOD / ShiLtJだけだということである。
ここが一番ややこしてところであるが、ミトコンドリアDNAと言うのは呼吸に関係しているから酸素といつも接触していて活性酸素の影響で突然変異が核DNAより頻繁に起きる。5倍から10倍の速度で変異が入る。
ところが、核DNAは細胞の中に1セットしかないが、ミトコンドリアは細胞の中に平均数千個ある。組織細胞の違いによって数も違う。卵の中だと24万個あると言われている。mtDNAは一個のミトコンドリアの中に十数個ある。人間の細胞は数十兆個あるとされているが、その数千倍のmtDNAの数の中で、一つ変異するのである。違う細胞内のどこかでは違う場所に一個の変異がはいる。或いは同じ細胞内の別のミトコンドリアには違う変異が入る。同じmtDNAの違う箇所に又別の変異が入る。
少々変異が入っても生きている限り何も問題ない。特にミトコンドリア病とされるまで特殊なケースだけが医学で問題とされる。普通の大多数の人の大部分の人生でミトコンドリアの突然変異は無関係である。寿命には何か関係があるとはされているが、寿命は何時かは誰にでも来る。
Ts.Markerさんの報告している現象はそういう普通の変異ではないということに気づかないといけない。変異は1つしかない。1つの変異だけが培養細胞の中で何割という増え方をしているのである。
体細胞に系譜決定されてしまった細胞内では変異は拡大しない。10個の細胞中に一つだけ変異が入っている細胞があるとすると確率的には倍に増えると2個/20個になるだけです。死滅細胞が補充されるときそういう確率で補充される。
一個でも変異ntDNAの入っている細胞の状態をヘテロプラスミーと呼んでいる。変異の入り方はどういう入り方でもいい。細胞内の全てのntDNAに他と違う異質の変異DNAがある状態をヘテロプラスミーと呼ぶ。何か所も変異していていいし、何個も変異ミトコンドリアがあっていい。そして年を取るほどそういう細胞は多くなる。そして全然問題ない。
問題は卵細胞です。卵は1個で独立している細胞なんですね。この1個から数十兆個の細胞に分裂分化増殖していく。従って、この卵の中のミトコンドリアは正常なmtDNAだけを持っているのが理想です。このすべてが同じntDNAで揃っている細胞の状態をホモプラスミーと呼んでいる。
この卵細胞は卵割のかなり初期に将来の卵細胞になる場所が決まっている。そして卵原細胞から卵になるときにそれがヘテロだったら一度ボトルネック効果を使ってホモに戻され、排卵までの待機中に維持されていて、ここで変異が入るとどうなるのかもよくわかって無くて、受精後胚盤胞になる間にもまだボトルネック効果が使われているらしい。
(
どうなってんの?)分からんもの同士で何とか検討しようということです。
まずボトルネック効果の仕組みが分かってない。昔はいくつか変異の入っているミトコンドリアがあるとまず総数を極度に減らして変異ミトコンドリアを消してしまう。そして残った正常ミトコンドリアを増殖させるという仮説だったが、調べてみると数は数百程度にまでしか減っていないと分かった。それだけでは十分に排除できないので、核の側から何らかの指示が出ていて、ヘテロプラスミーミトコンドリアを排除するようになっているのではとも言われている。
逆にボトルネック効果で変異が増えてしまった卵はそれが致命的な変異である場合は卵細胞ごとアポトーシスするということである。
今回Ts.Marker氏が紹介してくれている論文は、受精後の段階でもまだボトルネック効果で、変異ミトコンドリアを含んでいる細胞を隔離しようとする働きがあるという証明になっている。これは生まれて後に死に至ることによって次世代への影響を排除していることになる。また、その効果はES細胞、つまりインナーセルマス段階でも残っているようである。ES細胞化した時にまだボトルネック効果が働いていて培養中の継代で更に隔離されていくのかどうかまでは分かってない。
和モガ氏はES段階になってもまだ培養中のボルネックがあるという説になっているが、そもそもその仕組みは分かってない。
>>
しかし、いったん出来たSTAP幹細胞を培養するとSTAP幹細胞は増殖するので、ミトコンドリアがホモプラスミー化することになる。専門家ですら確かには分かっていないことをど素人がどれだけ検討できるかということである。
しかし、我々の目的はなぜそうなっているのかを事件の視点から解明するだけであるから、専門家のようにとことん分からなければ、分かったことにならないということではない。どこまでそれが可能か、考えてみるより仕様がない。なぜ、一塩基だけ違うヘテロプラスミーが全体の何割というオーダーで存在し得るのか。
(細胞株の混合)一番考えやすいのは二種類の幹細胞が混ぜられているケースである。この特殊な変異は一か所だけでしか起きていない。裏返すとTのホモプラスミーの細胞とAのホモプラスミーの細胞を混合して、それをPCRに掛けると混合割合が分かるというケースである。
この場合、ボトルネック効果などは何の関係もない。
問題はTであるマウス種が大半で、この場所がAであることが当たり前のマウスは今のところTs.Marker さんによってはNOD / ShiLtJしか見つけられていない。
他のマウスでたまたまこの場所がAに突然変異したmtDNAができたのだとしたら、今度はこの突然変異が卵細胞の全体を占めた原因に遡らなければならなくなる。この時は二つの幹細胞が混ぜられたという話では収まらなくなる。
まず最初に混ぜ合わせただけだという説から検討しよう。
この場合は割合は作成の時系列は無関係で、混ぜられた時の割合がたまたまそうなっていたというだけのことで単純である。単純でないのは細胞質の違う幹細胞が混ぜられたのはいいが、その両者の核DNAの変異特徴が一致しているということである。これを同時に説明できないといけない。
(FES1)
WGS DRR028646 FES1 T:3625(80%) A
:921(20%)(FES2)
WGS DRR028652 FES2 T:5307(100%) A:4
(ntESG1)-mtDNA=ICR
WGS DRR028644 ntESG1 T:4990(100%) A:4
(ntESG2)-mtDNA=ICR
WGS DRR028650 ntESG2 T:4291(100%) A:1
(CD45+)-mtDNA=129X1/Sv
Tru-seq SRR1171556 CD45+ T:2000(100%) A:1
(STAP)-mtDNA=129X1/Sv
Tru-seq SRR1171580 STAP T:2396(100%) A:1
ChIP-seq input SRR1171584 STAP T: 178(100%) A:0
SMARTer-seq SRR1171578 STAP T: 970(100%) A:1
(STAP-SC)-mtDNA=129X1/Sv
Tru-seq SRR1171585 STAP-SC(FLS?Acr)
T:1245(85%) A:217(15%) ??ChIP-seq input SRR1171589 STAP-SC T: 171(100%) A:0
WGS DRR028632 FLS3
T:1949(49%) A:1992(50%)WGS DRR028633 FLS3
T:1912(49%) A:2015(51%)(FI-SC)
Tru-seq SRR1171565 FI-SC(B6+α?) T:2212(100%) A:2
ChIP-seq input SRR1171569 FI-SC(CTS?Acr) T: 144(100%) A:0
WGS DRR028642 CTS1 T:3718(100%) A:4
WGS DRR028643 CTS1 T:3667(100%) A:5
(ESC)
Tru-seq SRR1171560 ESC T:1572(100%) A:0
ChIP-seq input SRR1171564 ESC T: 191(100%) A:0
SMARTer-seq SRR1171574 ESC T:1396(100%) A:0
(TSC)-mtDNA=ICR
Tru-seq SRR1171590 TSC T:2359(100%) A:0
ChIP-seq input SRR1171594 TSC T: 229(100%) A:0
(Epi-SC)-mtDNA=129X1/Sv
Tru-seq SRR1171558 Epi-SC T:1103(100%) A:0
(129/GFP ES)
WGS DRR028638 129/GFP ES
T:2963(80%) A:733(20%)(129CAGマウス)
WGS DRR028656 129CAG T:1219(100%) A:0
(129X1/SvJ マウス)
WGS Illumina HiSeq 2000 SRR1614029 129X1/SvJ T:3(100%) A:0 G:0 C:0
(ICR ?マウス )
SRR2917581 ICR ? T:219(100%) A:0
(DBA/2Jマウス)
Mouse strain assembly hub DBA/2J T
(129S1/SvImJマウス)
Mouse strain assembly hub 129S1/SvImJ T
(BALB/cJマウス)
Mouse strain assembly hub BALB/cJ T
(C57BL/6NJマウス)
Mouse strain assembly hub C57BL/6NJ T
(NOD/ShiLtJマウス)
Mouse strain assembly hub NOD/ShiLtJ
AこうやってみるとFES1、FLS3、129/GFP ESの三つが混ぜ物だということだね。
和モガさんの最初の直感が正しそうだ。でも、129X1/SvはTだから、ホモでAがあり得る集団としてはICRとNOD/ShiLtJしかない。従って、混ぜられた細胞が受精卵ESだということはあり得なくて、ntESだということになる。リシピエントマウスとしてAであるICRマウスか、NOD/ShiLtJが使われたとするよりないが、NOD/ShiLtJは免疫不全マウスなので通常はクローン胚のリシピエントとしては使われない。ICRマウスは近交系マウスではない。従ってAのICRマウスがあるのならば解決する。
もう一つの問題は何のために混ぜ合わせたのか、どうやって混ぜ合わせたのかということである。今まではすべてFLS3だと見ていた。これがntESだっただけで、若山さんが太田ESを取り寄せてFES1にFLSを混ぜたとした。129/GFP ESも同様にFES1にFLS3を入れたものだとしていた。ところが、このミトコンドリアDNAの変異分布がその可能性を許さなくなった。
我々の推測は若山さんがFLS3を作るとき岡部マウスのF1を小保方さんに渡していて、その酸浴細胞核を使ってntESにしたのだとした。だから捏造疑惑から逃れるために、小保方さんが太田ESをどこかで見つけて自分に渡したのだというストーリーにした。だから太田さんから取り寄せたFES1にはFLS3を入れ、学生のGOF-ESにはGLSを入れたのだ。そしてF1に関しては「僕のマウス」を渡したのだと嘘をついたと考えた。
しかし、ここで、若山さんはFLS3をそのまま入れたのではなくて、FLS3もAがリシピエントの別のntESと混ぜている。なぜかということである。
FLSは1-8まである。1は2012/1/31培養開始、2-8は2012/2/2培養開始である。FLS3はワンノブゼムである。ミトコンドリアDNAに関しては3しか調べられていない。そもそもなぜFLS3が全解析を受けたのか。
実はFLS3が全解析されることになったきっかけは最初に丹羽さんが2014/4/17にそれを持ち出して解析したからである。これは若山さんは関与していない。丹羽さんが偶然にこれを選んだだけである。
引っ越しの時に全ライン入れ替えてることになる。どうしてこんなことをしなければならないか。FLSは全部小保方さんがFES1を自分に渡したからだというストーリーにするのはFES1の中身をFLS3に入れ替えるだけでいい。FLS1-8までどうして全部入れ替えなければならないのか。
まして小保方さんはFLSとGLSに関しては自分の株分けした別ラインも持っていて、木星リストに書かれているが、ちゃんと書かれていなくてどこにそれがあるやもしれない。入れ替え犯が一番楽なのは大本を入れ替えてしまうことである。普通はそうする。
だからやはり入れ替えは普通にそうされたのだと仮定する。では、ヘテロプラスミーは最初から5対5でFLS3にあったのだ。
その場合はこのヘテロプラスミーは細胞が単に半々に混ぜられたからではない。
では生殖細胞が作られるときや、Ts.Marker さんが主張している第二次ボトルネック効果によって、Aという偶然にできた卵細胞の変異が卵や初期胚内にコンデンスされてそういう割合になっていたのだということになる。このメカニズムは専門家でもまだ全く分かって無くて、いろんな仮説検証をしている段階です。根拠が弱い。
なんて論理なんだと既知の知識内で専門家の間違いを正すなんてことは簡単で、それは専門領域の話で無くて、一般的良識の問題です。そうでないと裁判官は判決を書けないでしょう。でも、今遺跡を掘っている考古学者に先んじて、何が出るかを言えるど素人はいません。それを専門的領域という。ヘテロプラスミーの問題はまさに今専門家が遺跡にトレンチを入れている状況なんですね。ど素人が固唾をのんで見守っている。
まあ、我々はその道の領域に触れないままでこの事件の解に至れるかどうかを確認するだけのことしかできない。運よくそうであったらこの事件は解決するでしょう。別にヘテロプラスミーの深遠な科学的事実解明がどうなるかとは無関係である。
まず事実がある。FLS3が遺伝子全解析された2014年の7月以降から11月に頃に掛けて、解析サンプルのmtDNAの12188番地のTであるべきローカスにAという変異の有るmtDNAの割合が全体の半分を占めていた。この事実の可能的時系列変化の全てを辿れるかという問題である。まず素人でも分かることはこの変異が16万か所ある中の特定の1か所だけだということだ。他の場所は関係していない。この変異は時系列のどこかでたった一つのミトコンドリア内のたった一個のmtDNAのたった一か所のローカスで起きた。
変異の無いmtDNAとは何か。半分を占めているAでないTのmtDNAである。つまり変異しているAがTであったらすべてのmtDNAは正常なのである。
NOD/ShiLtJではこの場所はAであるのが正常である。Aであるからと言って何か機能障害があるわけではない。その証拠に近交系マウスとして長く維持されている。
今、FLS3に入っているAはマウス系統としてどこかから入り込んだのではないという前提で考えて居る。どこかから入り込んだものが混ぜられた結果半々になったということは無さそうだという結論から、今、突然変異が排除されずに残ったのだという前提で考え直しているのである。核酸の種類は4つしかないのだから変異が偶然Aになることも不思議でもなんでもない。
この変異がいつ入ったのかは分からないが、胎児段階以降に入ったと仮定するとどうなるか。ある細胞にこの変異したミトコンドリアが存在する。これが二つに分裂したときどちらかに変異したミトコンドリアが入る。一方は正常で他方は変異ミトコンドリアを1個含む。細胞質分割をしているから細胞分裂後にどちらの細胞も細胞質を倍に復元する。その時に変異ミトコンドリアを含んでいる方は二個に増える。全体では1個だった細胞が2個に増え、変異している方の変異ミトコンドリア数は2個になる。更に細胞分裂するとどうなるかというと、正常な方の細胞1個は二つに分かれて2個になる。変異しているミトコンドリアを2個含んでいる細胞も二つに分かれるが、この時に二個とも一方に偏ると、半分は正常、もう一つは4個の変異ミトコンドリアを持つ変異細胞になる。つまり、全体では4個の細胞になり、内3個は正常、1個は変異細胞である。これを繰り返すと変異細胞の中の変異ミトコンドリア数は増えていくが、全体の細胞数の下で、この細胞を拾い出す確率はどんどん下がるということが分かる。
他方、2個の変異ミトコンドリアを持つ細胞が一個ずつに分配されて分裂した時はどうなるかというと、全体では4個の細胞は同じであるが、二個が正常で残りの二個の変異細胞はそれぞれ変異ミトコンドリアを2個持つ細胞になる。変異したミトコンドリアの数が全体では4個なのは変わらないことは注意しておくべきである。そして胎児内のこの変異ミトコンドリア数は細胞分裂の度に2のn乗の関数で増えていく。
生体内ではこの変異がいつの時点で発生したかは重要である。というのも胎児の細胞数が相当程度に大きくなっている段階で1個の変異が起きてもこのメカニズムでは全体の中のその時点での割合を超えるということはない。むしろコンデンスされるケースでは割合は減る。
ではこの極端な例として1個の細胞時、つまり卵の中に1個の変異ミトコンドリアがあった場合を考える。この場合は普通の細胞分裂だけを考えると、半分に分割されたときにどちらかに変異ミトコンドリアが入るから、この時点で正常細胞と変異細胞に分かれ、変異細胞内の変異ミトコンドリアは2個になる。ここから細胞分裂を繰り返すと、最大変異細胞数は全体の半分になることが可能で、たぶんそれ以下になることは可能であるが、それ以上には決してなれない。
FLS3が半々であることの原因に近づいてはいるが、まだ十分ではない。なぜなら今はマウス体内での話をしていて、人工的な培養の話ではないからである。
ここまで、そもそもAであることは何の機能障害もないのであるから、成体の生存には関係しない。その証拠にNOD/ShiLtJはAである。ここで上の極端な事例と勘違いしないようにしないといけないが、NOD/ShiLtJはそもそも卵の中の全てのミトコンドリア、或はその中に十数個あるmtDNAの一か所がAなのだということです。どこまで分割されても100%Aになる。これがTと半々になるということがあるとしたら、別のマウス系統の細胞質と混ぜられているからだ。でも、FES3は若山さんは何か別の系統を二つ混ぜたわけではなさそうだということで、今検討している。結構ややこしいので混同しないようにしないといけない。
FLS3の1個の変異がどこで発生したかはまだまだ全然わかっていない。まず胎児段階のどこかで、つまりボトルネック効果の働かない時点での胎児の場合に発生していたらどういうことになるのかという可能性検討で、この変異の広がりの限界が最初の胎児の細胞数との比率を超えないということと、コンデンスされても程度があるということが分かった。なぜこの成体の中でどういう広がり方をするかを確認しておかなければならないかというと、次世代の卵が作られるときにその卵の場所にこの変異細胞がある確率も考えないといけないからである。
実は卵原細胞が出来る場所が決定されるのはとても早い時期で、卵の分割が起こるときに例えば8細胞期に上半分には絶対存在しない。分割が進むにつれてその場所は局所化されていく。例えば、胎児が成体に近くなって心臓にこの変異が入ったとして、それが次世代に引き継がれるということは絶対にないと分かる。心臓があるような場所には卵原細胞はできないのである。心臓近辺に上述したような変異がいくら広がって行っても、その場所が卵細胞になることはない。この変異が卵細胞の直ぐそばで起こる確率がどの程度かはFLS3の半々結果の原因に直結しているのである。
さて、では、FLSはどうやって作られたか。ここにいろんな操作が疑われていて誰の言ってることが正しいのか全く調査されていないから困っているわけである。警察を入れていればこんなところは既に事実関係として確定されているでしょう。でも桂報告は個人の恣意判断だけで推測を書いているだけだ。
科学的解析結果として分かっているのは、核は129/B6-Acr-CAGで細胞自体にはY染色体が存在していて全ライン雄の細胞だとされている。このFLS由来4Nキメラは全部雄なのでジャームライントランスミッション確認するときには兄妹交配ができないので、別の雌をもって来なければならなかった。GFPが半分しかこなかった以前に雌のキメラが居なかったから別の雌を持ってきたんです。この時点で変だと思わない方がどうかしている。自分でわざわざ129の親メスを使って交配実験している。嘘ついてるんです。
渡した赤ちゃんマウスは雌雄取り混ぜている。そのことは論文に書いてあるし、かつ若山さんの記者会見でも本人がそういってる。ところが4Nキメラは全部雄だった。
論文通りに塊でSTAP細胞を培養誘導してSTAP幹細胞を作ったのだとしたら、その細胞は雌雄を半々に含む。それを4Nキメラ胚に入れたら、雌雄半々で入っていますから偶然に生殖細胞に当たったSTAP幹細胞の雌雄確率に従って、つまり、半々に雌雄が生まれるはずですが、8株全部に平均30個もキメラ胚を作って、生まれて成長したキメラトータル26匹全部雄だった。これを不思議にも思わずに、雌が居ないから兄妹交配できないとして、129のメス親と交配した。この時点で既にそれを不思議と思ってないことが分かる。GFPが半分にしか来なかったなんて言い出したのは2012春にヴァカンティが米国特許仮申請を始めようとしていた時期です。そしてそれ以降も実験をそのまま続けている。犯人が誰かなんて我々にはとっくに分かっていることです。繰り返しません。
桂報告書ではこれが全部雄だったのは雄であった大田ESを小保方さんがコンタミさせたからだということになっているわけです。でもこれは何の根拠もない。でも、事実として、FLS1-8は全株雄なんですね。なぜそうなるのか。
論文通りならまずSTAP細胞自体が雌雄半々の細胞集団のはずです。そしてSTAPキメラは作られている。これが既に幹細胞がどうこう言う以前に4Nキメラでも雌雄半々に生まれていないといけない。そして4Nキメラの実験は最初から行われていて、2011/11/28のキメラ写真は4Nキメラでしたね。
ところがジャームライントランスミッション結果は2Nキメラの分しかありません。そしてこれは兄妹交配していますが、2Nだとリシピエントの細胞が生殖細胞になっていると雌も生まれ得るから、ここは4Nキメラのデータがないといけない。ところがそのデータはアーティクルではつけられていないのです。
これは若山さんの意図的な選択です。STAP由来4Nキメラも雄ばかりだったからです。
なぜそうなるのか。
小保方さんは成功したキメラ実験の前にF1のSTAP細胞を渡しています。そしてこれはキメラ成功していない。次にもう一度渡したときにナイフ切り分けで成功したことになった。
この成功した細胞を渡されたときからクローン胚を作っていると10日後のキメラ成功には間に合いません。前回に失敗した時の分を使って同時にクローン胚を作っている。クローンの成功率は低い。たくさん作っています。小保方さんはこの細胞ではキメラはできていませんから、太田細胞なんて入っていない。
クローン胚に入れられた細胞は雌雄半々だったんです。成功率1%です。この中にもいくつかの成功細胞があるので半々に雌雄があったでしょう。そしてこの胚盤胞からESを何ラインか作った。そしてそれぞれのラインを4Nキメラ胚に入れて樹立確認をした。ここでも成功率は2/3でしょうか。うまく成功したラインが雄だけだったということです。
この原ntESが後にSTAP細胞と入れ替えられていくものです。小保方さんの作ったSTAP細胞は廃棄されて、小保方細胞核使用ntES由来のキメラが作られていく。
STAP細胞とSTAP幹細胞は若山さんにとっては同じものです。
小保方さんが自分で作って遺伝子解析に出しているSTAP細胞は若山さんのキメラのできるSTAP細胞=STAP幹細胞とは別ものです。
FLSの核は129/B6-Acr-CAGで細胞自体にはY染色体が存在していて全ライン雄だということは事実ですね。そしてどうしてそうなるかの一つのストーリー説明もした。
仮にntESだったとしたらどうなるかというと、ICRのリシピエント卵の中にこの変異ミトコンドリアがあった場合と、129のドナー卵のミトコンドリアが移植時に持つ込まれた影響と両方考えることになる。
一応、正常な状態ではどちらもTなんですからAはいずれにせよ突然変異で持ち込まれたものということになる。
まずICRの卵の中に既にこのヘテロプラスミーがあった場合です。
動物実験申請書ではICRはマウス業者から購入されている。自家繁殖ではないようです。
流石にマウス業者から買った時はmtDNAは登録データ通りにホモプラスミーで販売されているのではないでしょうか。ここも素人には分からないところです。そんなところまで管理しているのかどうか。でも、管理していなければ登録データには載せられませんからね。そう推測するわけです。
買ったときにはホモプラスミーであったのなら、変異は2011年の実験時以降に発生している。このICRマウスの1個の変異が全体の50%になるということはありませんね。ここでは生殖細胞ボトルネックは経過していない。一度もマウスには戻っていません。クローン胚の中で胚盤胞になり、一度ESになって、更にキメラ胚の中で確認はされたが残っている細胞はそのまま培養されている。従って、この時の変異は培養中に全体のmtDNA総量の50%まで増えたことになる。
正常な細胞が存在している中で無限増殖していくと細胞数で最初の比率のまま増えていくことはあり得ますが、mtDNA総量が半分にまで増えるということはあり得ません。他の細胞の中の正常なmtDNAも同率で増えていく上に、変異ミトコンドリア含む細胞に正常なミトコンドリアが圧倒的に存在している。
この可能性はない。
では、129のドナー側からクローン胚にもたらされたらどうか。この場合でも実験以降にAが入ったとしたら同じです。違うのは、この実験時点で既に129側のヘテロプラスミー率が高かった場合です。マウスで継代されるとヘテロプラスミー率はコンデンスされて高まる卵が出てくる。無論病的な変異であれば卵ごとアポトーシスされるが、このAの一点だけの変異は無害ですからアポトーシス排除はされない。
でも、クローン胚に入れられた時にごくわずかしか入ってないですから、この時点で、一個の卵の中での変異ミトコンドリアの比率は格段に減っている。これが二分割されると一個は正常他方は変異細胞。正常側は常に半分存在していて、残りの変異側から正常細胞が分離されていくので、全体の変異ミトコンドリア数は半分まではとてもいけない。
では太田細胞だったのであればどうなるのか。これはまず太田細胞が何なのか全く厳密な調査を受けていないから分からない。太田さんの言ったとおりに作られていたらterなんでしょ。でもX1だったのならそうでしょうなんて言う証言では何が本当かを確定させて論じることができない。ましてなかったはずだがあったのなら置き忘れでしょうなんて、何を言ってるんだ。国民をバカにしてるのかというレヴェルです。お話にならない。
でも、まあ129/B6-Acr-CAGだったとしましょう。しかも珍しく受精卵ESを目的も無く作った。そして使いもせずにすぐに凍結した。凍結したら細胞は遺伝的に変化しませんからね。ミトコンドリアも固まってじっとしています。で、小保方さんが2011年の11月頃に解凍したということですね。細胞質は129X1ですね。ミトコンドリアのその場所はTが普通だ。この129はどこから持ってきたものなんですかね。市販のものだったらTのホモプラスミーでしょうね。変異は実験後に入ったということですね。一度もマウスにはなってませんね。ESのまま凍結されていた。これが20%の割合にまで増えることはありません。従って、市販のマウスではなく、若山研で自家飼育されていた129で既にヘテロプラスミー割合が高まっていたものということになる。
ところがFES1は2割ですが、同じコロニーの129マウスであるはずのFES2はゼロです。
いろんな可能性を考える以前に人の嘘を確認する方が先でしょうね。これは何らかの人為です。
そしていよいよ、今度は胎児段階ではなく、卵から胚盤胞期、そしてES段階で変異が入った場合です。私の基礎知識は林純一氏の『ミトコンドリア・ミステリー』です。初版が2002/11/20で、現在からみて18年前です。この頃以前に精子のミトコンドリアが遺伝するという説もあって、長い検討の末、精子のミトコンドリアは殺されるということが分かった。この人の功績で、以後、定説は動いていない。ブルーバックスシリーズですから一般教養です。2002年ですらもう一応定説になったことが基本書かれている。発見はもう少し前なんですね。
20年以上経過している。今度Ts.Markerさんが紹介してくれた論文は、Rapid mitochondrial DNA segregation in primate preimplantation embryos precedes somatic and germline bottleneckというタイトルで、2012/5/3がオンライン発行日です。林さんの本からは10年経過している。そしてこの論文自体が更にそこから8年経過している。
ど素人なので今どういう評価なのかは分からない。
参考までにもう一つ「マウスにおけるmtDNA遺伝様式 : 母性遺伝と急調分離」という日本語論文がある。
© 2011 The Japanese Society of Plant Morphology とあって、これもやはり8年以上前である。興味深いのは著者で、設楽浩志, 米川博通氏の共著だが、米川さんは桂報告書に名を連ねているあの人ですね。

ボトルネックのメカニズムそのものが見直されようとしている。
>>
mtDNAコピー数の極端な減少が, ボトルネック効果の原因ではなかったことから, 他のメカニズムによって ボトルネック効果が起こり, 急調分離を成立させている可能性が高い.
現在までに, 我々は2つの遺伝モデ ルを提唱している(Cao et al. 2007). 一つは, 細胞中 のごく限られたmtDNAの集団が選択的に複製される としたモデルである(図3B). 複製されうるmtDNA分 子種が限定されることで, 異質性の割合が規定され, そのまま子孫に伝達された結果, 急調分離が引き起 こされることになる. 実際に前述のカナダのグループ は, 卵の成長過程において一部のmtDNAの集団が 複製されうることを観察し, この遺伝モデルによって急 調分離を引き起こされていることを示している(Wai et al. 2008).
もう一方のモデルが, mtDNAがサブグループ を形成することで, 分離単位の実効数を減少させるモ デルである(図3C). mtDNAはミトコンドリア内でミトコ ンドリア転写因子A(TFAM)やポリメラーゼγ(POLG) などと結合し, DNAとタンパクの複合体(核様体, ヌク レオイド)を形成している(Alam et al. 2003, Bogenhagen et al. 2008). このミトコンドリア核様体には, 2-10分 子のmtDNAが存在しているとされており(Legros et al. 2004), これがサブグループとしての機能を果たしうる. さらに, 哺乳類のミトコンドリアは融合, 分裂, 輸送を 繰り返し(Hirokawa 1998, Ishihara et al. 2003), ミトコン ドリア内の生理活性物質を混合することで, 細胞として はミトコンドリア内の構成成分の均一化されていると考え られているが, ミトコンドリア核様体間でのmtDNAが混合する頻度は比較的少ないことが示されており(Gilkerson et al. 2008), こうしたミトコンドリア動態からもこのモデル の可能性が十分に高いと考えられる.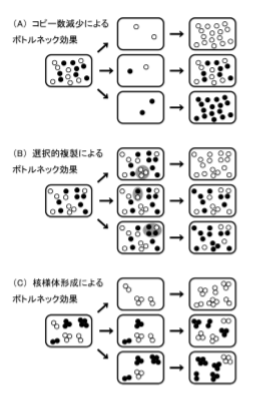
これも8年も経過しているんだから少しは進展して真偽が明らかになっていそうなものですが、何せ素人レヴェルでは分かりませんね。何事も素人は10年遅れですよね。
ともあれ、Ts.Marker さんの紹介論文はES段階でもまだボトルネック効果が発揮されているというんですから重大です。ちょっと検討してみましょう。
>>
Rapid mitochondrial DNA segregation in primate preimplantation embryos precedes somatic and germline bottleneck
Hyo-Sang Lee,1 Hong Ma,1 Rita Cervera Juanes,1 Masahito Tachibana,1 Michelle Sparman,1 Joy Woodward,1 Cathy Ramsey,1 Jing Xu,1 Eun-Ju Kang,1 Paula Amato,2 Georg Mair,3 Ralf Steinborn,3 and Shoukhrat Mitalipov1,2,4,5,*
SUMMARY
The timing and mechanisms of mitochondrial DNA (mtDNA) segregation and transmission in mammals are poorly understood. Genetic bottleneck in female germ cells has been proposed as the main phenomenon responsible for rapid intergenerational segregation of heteroplasmic mtDNA. We demonstrate here that mtDNA segregation occurs during primate preimplantation embryogenesis resulting in partitioning of mtDNA variants between daughter blastomeres. A substantial shift toward homoplasmy occurred in fetuses and embryonic stem cells (ESCs) derived from these heteroplasmic embryos. We also observed a wide range of heteroplasmic mtDNA variants distributed in individual oocytes recovered from these fetuses. Thus, we present here evidence for a previously unknown mtDNA segregation and bottleneck during preimplantation embryo development, suggesting that return to the homoplasmic condition can occur during development of an individual organism from the zygote to birth, without a passage through the germline.米川さんの論文は林さんの教科書的記載に対して、まずボトルネックはそんなに数は減って無いと主張した。だからメカニズムがどうも教科書的知見と違うと指摘している。対して、Ts.Markerさん紹介論文では、ボトルネック効果の起こる時期が違うと言ってる。米川さんのところの日本語用語では急調分離ですか。英語ではRapid segregation となってるから、どちらの論文も同じ現象を言ってる。前者はメカニズム、後者は時期ですね。
Figure1です。
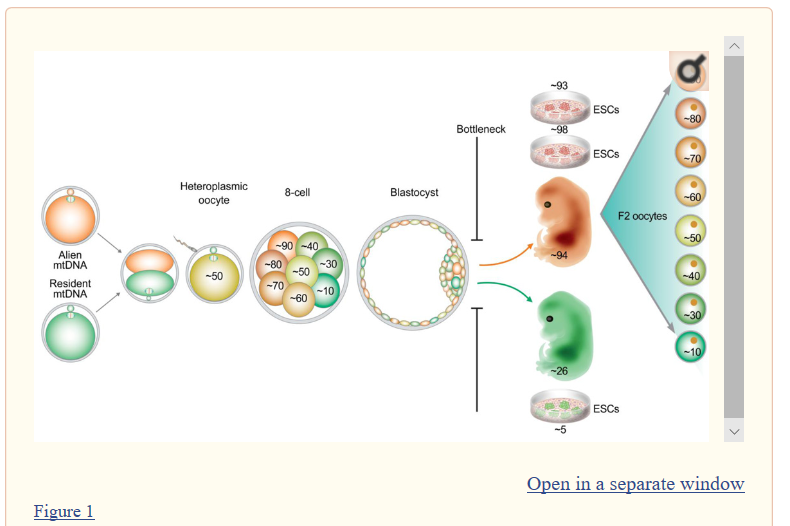
ボトルネックと言うのは卵母細胞の時点で起こるとされていたのを否定した。まず緑色の既にできている卵に対して細胞質を半分取り除いて別の種類の細胞質を注入する。一個の卵の中に二種類のmtDNAが入るのでヘテロプラスミーになる。PCRにかけてほぼ半々であることを確認している。
これと似たようなPCR結果が、上述している、二種類のサンプルを混ぜたケースである。違いは1個の細胞か、たくさんの細胞かの違いだけである。
ともあれ、この実験では卵母細胞の発生時期にのみボトルネックが起きるのではないということの1ケースを実証した。図では卵の大きさは8細胞期で拡大されているが、胚盤胞期までほとんど同じ大きさである。ハッチング前に卵の外側の膜が柔らかくなって水分を吸い取ってやや大きくなった後に殻を脱ぎ捨てて着床する。
8細胞期なんてそもそも大きさは変わっていない。中を分かりやすくするために拡大表示しているだけである。ここは勘違いしないようにしなければならない。分割球の一個ずつはどんどん小さく分割されていくのである。つまり胎児段階の体細胞分裂と違っては半分に細胞質がなったあとに倍には戻らないで、そのままどんどん容積が小さくなっていくだけである。
従って胚盤胞期の各細胞は小さすぎで個別解析はされていない。出来なかったと書いてある。8細胞期までを調べたのである。
その結果8細胞期の図にあるように、8個の細胞はヘテロプラスミー率がすべて50%に均等に分裂したかというと、そうではなくて全部がホモになろうと努力していたことが分かったということである。この胚盤胞期に至る発生期においてもボトルネックによる急調分離が起きているという証明である。
しかもこの段階ではミトコンドリア数は昔謂われていたように数は減っていない。ランダム選択説はこの発生期の急調分離メカニズムとしては通用していないという。つまり米川さんの論文と同じことを指摘しているようだ。要するにボトルネックのメカニズムはまだ分かってないということで、ここでは何か別のメカニズムがあると示唆している。
でも、要する分かってないということです。プロがまだ分かってないことに素人があれこれ論じても意味がないですね。せいぜい、発生期でもまだmtDNAはホモプラスミーに揃おうとしているらしいということを確認できればいいのではないか。つまり分かってないところは断定するなということです。
ES細胞になってから培養中にまだホモプラスミーに揃おうとしているのであろうか。分かっていないということです。
(Ts.Markerさんの問い)以下、Ts.Markerさんの疑問であった。
>>
heteroplasmy の違いから - ni -
2020年1月24日
ミトコンドリア病(ヘテロプラスミー)についての情報は近年かなり豊富なようだ。暇に任せ調べていくうちにヘテロプラスミーの比率を変化させるもう一つの知見があった!
それは卵子形成時のボトルネック効果によるヘテロプラスミー割合の変動のみならず胚盤胞(Blastocyst)期でもボトルネック効果があるという。
卵子形成時の早期ボトルネックに対し胚盤胞期のボトルネックは後期ボトルネック(second bottleneack)と呼ばれヘテロプラスミー比率の変動が報告されている。
初期の論文「Rapid Mitochondrial DNA Segregation in Primate Preimplantation Embryos Precedes Somatic and Germline Bottleneck」
イメージ(https://www.lifescience.mext.go.jp/files/pdf/n2132_06.pdf#search=%27blastocyst+bottleneck%27)
例えば、ヘテロプラスミーをもつFES1の株分けらしい129/GFP ESを4倍体胚盤胞にインジェクションし4Nキメラマウスを得る。その4Nキメラマウスを筆頭著者に渡していたらsecond bottleneckによりヘテロプラスミーの比率はばらつきFES1,129/GFP ESとは違ってくるだろうということは容易に想像できる。Aの比率が増えたり(FLS3) Tの比率が増えたり(FLS1) ホモプラスミーになったり(CTS1)...。
エアF1マウスを渡されていたんじゃないか...。①例えば、ヘテロプラスミーをもつFES1の株分けらしい129/GFP ESをFES1の中身が太田さんの作ったものか否かは問わず、桂報告書が分析したFES1という意味で、129/GFP ESはその株分けだと。
②4倍体胚盤胞にインジェクションし4Nキメラマウスを得る。若山さんがなぜそんなことを当時しなければならなかったのかの状況説明が何も書かれていないが、仮にそうしたとして、その129/GFP ESの4Nキメラは簡単に作れる。
③その4Nキメラマウスを筆頭著者に渡していたらsecond bottleneckによりヘテロプラスミーの比率はばらつきFES1,129/GFP ESとは違ってくるだろうということは容易に想像できる。その4Nキメラ胚にES細胞を入れたらsecond bottleneckが必ず起こるのなら、生まれてきた4Nキメラマウスはそれぞれすべてヘテロプラスミー率が違うであろうが、キメラ実験した結果は論文には書かれていないので推測に過ぎない。
④Aの比率が増えたり(FLS3) Tの比率が増えたり(FLS1) ホモプラスミーになったり(CTS1)...。
事件化する前に若山さんが太田ESとされている細胞及びその株分け細胞を使って4Nキメラを作っていて、この赤ちゃんマウスを渡して小保方さんに酸浴細胞を作らせていたが、成功せず、次にナイフ切り分けで、キメラができ、幹細胞かもできたと。若山さんは最初にどうしてそんな複雑なことをしなければならなかったのでしょうか。どうして岡部マウスと129のF1を渡していたと考えてはいけないのでしょうか。
それだと事件化後に発見されたmtDNAの変異箇所の説明がつかないからですね。でも、よく考えてください。FES1も129/GFP ESもラボにはなかったもので、小保方さんが置忘れを見つけたとされているのではないですか。それなら若山さんはそんな4Nキメラは作りようがない。
逆に若山さんがそのF1の細胞を持っていたのだとしましょう。そのような4Nキメラの赤ちゃんマウスを渡していたのだとしましょう。それでも小保方さんはそのマウスの体細胞を酸浴させてSTAP細胞を作って若山さんに手渡した。最初できなかったが、次にナイフで切り分けてできた。そして幹細胞もできた。これって大成功ではないですか。どうして若山さんが論文取り下げに躍起にならなければならないのでしょうか。これって元が何であれ、体細胞リプログラミングの証明です。太田ESから作られた4Nキメラマウスを使ったからと言って、実験結果に何の影響もない。
しかも、再現実験でキメラはできなかった。
⑤エアF1マウスを渡されていたんじゃないか...。(⑤エアF1マウスを渡されていたんじゃないか...。)Ts.Markerさんは"エアF1"という言葉を使う。
>>
0. Zscan4
2020年01月21日 14:40
>>8
>..の部分は中身の話ですね。
相変わらずかみ合わないね。
129/GFP ES を起こして、インジェクションし4Nキメラの129B6マウスを生ませる。(エアF1)まずエアの意味が分からない。
エアギターというのは本物のギターは無いのにそれを演奏している形態模写を言うから、エアF1というからには本物のF1ではなく、偽物のF1だという意味なのであろう。マウスを掛け合わせて生まれた子供がF1マウスだとするとそのESもF1ESなので、とても紛らわしい。しかし、常識的に解釈して、エアF1というのは生きているマウスなのであってESではないという意味なのは明確だ。従ってエアF1とTs.Marker さんが揶揄しているのは、マウスだと思わせといて実際には盲点になっているが、ES細胞を使ってるだろうという批判ですよね。
ここまではいいんですね。
問題は、どうしてES細胞だと思うのかというところです。ES細胞から作った4Nキメラじゃないかと考えるのは自由なので、その可能性を追うのは構わないが、そうであったら、この事件のどこがどう変わるのかの説明が全くないから理解できない。噛み合ってないのではなくて、説明を受けていないから分からないということですね。
どこかで、説明が書かれているのならそこを指定してくれたら読むが、一応Ts.Marker さんのブログは全部チェックしている。事件を概括している説明はないはずである。この辺りは和モガさんと対照的だ。しかし、Ts.Marker さんの示唆している推測できる概括は和モガさんのと違うようだ。
(推測できるTs.Marker さん説)誰でも知っている通り、Ts.Marker さんは和モガさんと並んで早くからの桂報告書の批判者です。どちらもどうやら理系の人らしい。どの程度このSTAP関連の専門に近いのかは分かってない。和モガさんはいち早くFES1とFLS3が=であるということはどちらが先であったか、つまり太田ESでFES3が捏造されたのか、逆にFLS3で太田ESが捏造されたのかの証明にはなっていないと指摘した。
Ts.Markerさんは、SNP解析が変だと指摘し、FI-SCのOct4-GFPは光ってるじゃないか等々と、最初から細かい疑念を提出した。私はど素人なので、桂報告を読んだ時は感心して、なんだこんなに細かく分析してるんなら、簡単に犯人は分かるはずだと思った。特に何度もES細胞でコンタミさせている結論になってるのに意図的コンタミであるかどうかも分からないなんて、頓珍漢な話なので、これは裏があつてわざと結論を曖昧にしているのだと直感した。その時はむしろ小保方さん犯人だろうと思っていた段階だった。
一般人としては、まずは何かの勘違いだ。次はNHKを見て中国人のスパイじゃないか。そして、次は若山さんの記者会見を見て、奥さんを庇ってるのかとも思ったが、桂報告最初に読んだ時は小保方さんだと分かったのだなと推測した。だからわざと結論をぼかしていると考えて、小保方さん犯人説を確定させてやろうと考えた。できなかった。どうしてもおかしい。そして手記が出た。ここにリクルートの問題が絡んでいたと知った。
①キメラは論文通りにできた。(和モガ、Ts.Marker説)
②キメラは小保方さんのESコンタミ捏造である。(桂報告書結論)
③キメラは小保方さんの成功を妬んだ第三者が以後妨害コンタミを続けた。(和モガ説)
④キメラは若山さんの小保方核使用ntESによって作られ、リクルート上の都合で一時的嘘をついたもの。(居士説)
⑤キメラはできていて、幹細胞化は共培養によるもの。(Ts.Marker 説)
和モガさんは①③
①キメラは論文通りにできた。(和モガ、Ts.Marker説)
③キメラは小保方さんの成功を妬んだ第三者が以後妨害コンタミを続けた。(和モガ説)
Ts.Markerさんは①⑤
①キメラは論文通りにできた。(和モガ、Ts.Marker説)
⑤キメラはできていて、幹細胞化は共培養によるもの。(Ts.Marker 説)
居士説は④
④キメラは若山さんの小保方核使用ntESによって作られ、リクルート上の都合で一時的嘘をついたもの。(居士説)
桂報告書説は②
②キメラは小保方さんのESコンタミ捏造である。(桂報告書結論)
STAPキメラに関して分類すると
A.キメラは論文通りできた。
和モガさん、Ts.Markerさん
B.キメラは論文通りにはできてない。
桂報告書、一言居士
STAP幹細胞に関して分類すると
A.幹細胞は論文通りにできた。
和モガさん
B.幹細胞は論文通りにはできてない。
桂報告書、Ts.Markerさん、一言居士
Ooboe さんから私の和モガ説批判を再掲してくれとの依頼がコメント欄にあるが、先にTs.Marker 説から検証する。(Ts.Marker説構築)和モガさんはこの事件をきれいなフローチャートに纏めてくれているので、推測の流れがとても良く分かって批判もしやすい。Ts.Marker さんは何か故あって正体を知られたくないのか、多くを語らない。ほとんど仄めかしなので、これでは第三者が理解するのは難しい。しかし、多く語ると文は人なりと言って人間性がでてしまうので、人それぞれ都合があろうから、それをとやかく言っても仕方ない。
しかし、「相変わらずかみ合わないね。」という言葉の裏には噛み合っ欲しいという希望は見えるので、推測してもらいたいという意味であろう。推測してこれでいいんですかと問うても答えないという方針なのは分かっているので、甲斐がないこと甚だしいが、ただ、言葉の端々を繋いで全体を構築しておいてみる程度のことは出来る。
今まで一番彼が自分の推測を語ったのは共培養の件である。
>>
23. Zscan4
2019年12月15日 21:23
>>22
論文通りでは、幹細胞は再現できなかったと筆頭著者さんが手記に書いている。
別のプラスアルファーがあったんじゃなかろうかと。
「ESやTSとの共培養による幹細胞への誘導」
あの日に書かれた3T3細胞のくだりを覚えている?
共培養により他の細胞からの因子を受けて増殖する話。
和モガさんも乗ってくれたが、他の方のアドバイスも頂いた。
ESやTSとの共培養は非常にリスキーだと(DORAのブログ参照 気まぐれ先生より)
幹細胞に駆逐され入れ替わる場合もということ。
また後のソートも難しい、10%ぐらいTS(CD1)が残ったというのも考えられるんじゃない。まず彼はキメラは論文通りできていると考えて居る。ただ、幹細胞は小保方さんに出来なかった。だからここで若山さんが論文に書かれていない手技を使ったのだ。それがESとの共培養だと考えて居る。TSのことはともかくとして、ここまでは最初のキメラ成功時に行われた結果を説明していることにならないといけない。
リスキーではあっても、成功したから幹細胞もできたと言った。ここで、彼がヘテロプラスミーに関して推測したことを加えると、最初に成功したキメラマウスの由来細胞は大田ESだということになる。
まず若山さんは後に桂報告書で太田ESとかFES1と呼ばれた細胞の元を持っているのでないといけない。次にこのES細胞を使って4Nキメラを作った。そして小保方さんに129B6F1マウスだと言ってその赤ちゃんマウスを渡した。
①129xB6-Acr-CAGF1ESから作られた4Nキメラ赤ちゃんマウスである。
②129xB6-Acr-CAGF1の自然発生マウスである。
どちらかということになるが、どちらであっても小保方さんはそれを酸浴させてSTAP細胞を作る。形態判断でそのスフィア塊を若山さんに渡す。若山さんはナイフカットして2Nと4Nキメラ胚にインジェクトする。数百個のインジェクションを行う。
キメラはそれでできた。4Nキメラの帝王切開胎児が2011/11/28に取り出された。4Nの方が良く光るので見栄えがいいから4Nが取り出された。2Nマウスは自然発生させたのであろうか。その場合は更に10日後の2011/12/10ごろになるが、論文にはこの写真はない。2011/11/25にはGLが樹立開始されている。キメラが成功しているか否かは4Nキメラのリシピエントのお腹が膨れてきたら2011/11/28以前に分かるので、GOFでも幹細胞化を試したのだと分かる。この時のGOFのキメラ実験が行なわれたのか否かも発表されていない。若山さんの実験ノートはすべて提出されていたはずである。しかし、GLの検査すらされていないのであるから、キメラに関しても触れられていない。GOFのキメラは作られていたとしたら、2011/11/25のキメラ胚から作るとされた幹細胞と同時期の異色のはずなので、帝王切開なら2011/12/5頃、自然出産は2011/12/15頃になる。因みに小保方さんは12/10から12/18まで渡米しているので自然出産に立ち会ってはいないことになる。
私はキメラができたのも幹細胞ができたのもntESだと思っているが、Ts.Marker さんはキメラはナイフ切り分けでできた。幹細胞は一旦キメラ胚の中に入れただけでリシピエントのインナーセルマスとの共培養になるのでそれでできたと言ってるのか、キメラ胚に入れた時にES細胞とSTAP細胞を両方入れたのか、或は、キメラ胚には入れずに単に培養皿の上で共培養したものをFACS選別したら増殖し始めたのか、といった、細かいことの説明が無いのではっきりしない。しかし、以下の記述は培養皿の上ということなのであろうか。
>>
ESやTSとの共培養は非常にリスキーだと(DORAのブログ参照 気まぐれ先生より)
幹細胞に駆逐され入れ替わる場合もということ。
また後のソートも難しい、10%ぐらいTS(CD1)が残ったというのも考えられるんじゃない。最後のソートに関してはGFPの有無でソートするので大して難しくないと思うが。
ともあれ、小保方さんはGOFの赤ちゃんマウスと12/27検収のヌードマウスを使って、休日出勤してテラトーマを作り、その後渡米した。そして翌年に帰ってきてそのテラトーマを切り出して3誌論文に使わずにいたら、後の事件化後の検査でアクロシンが出た。
私は若山さんが幹細胞と称するntESを上から注射したからだと説明しているが、Ts.Markerさんはその説明がない。彼の説では共培養でできた幹細胞を注射したと説明すると、私と同じ説明がつくが、私と彼の説の違いは、彼はキメラは論文通りに出来ていると考えて居る。ならばどうして若山さんは幹細胞を注射したのか。そのままで出来るかどうかまずは自分でも確認したいのではないか。
私の説はキメラはntES化したからできたのであるから、小保方さんの酸浴細胞でテラトーマを作っても簡単に出来る筈はない。酸浴でとても光るようになったから、ティシュー論文や博論とは違ってできがいいはず、ましてキメラもできていると小保方さんは期待している。リクルートのための時間稼ぎの嘘に過ぎないのだから、ちょいと注射したと解釈している。Ts.Marker さんの推測ではそこの説明がうまくいかないのではないか。
(Ooboe さん要望の和モガ説批判再掲)和モガさんの説明はとてもクリアなので理解しやすい。まずは有名なフローチャートを貼り付けましょう。
カテゴリーの「STAP細胞事件(90)」をクリックして下までスクロールして5回ページをめくると出てくる。
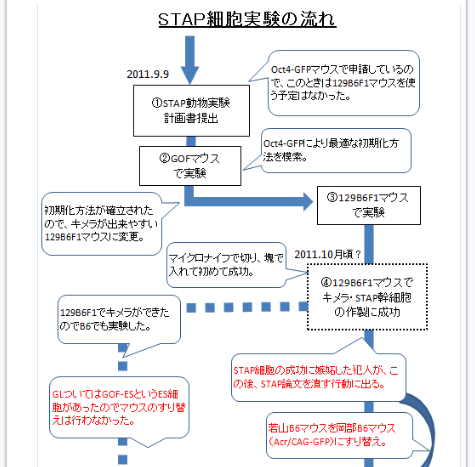
自分のブログを持つと図が使えるので説明しやすいですね。まず2011/9/9付の動物実験申請書は以下です。

途中省略します。
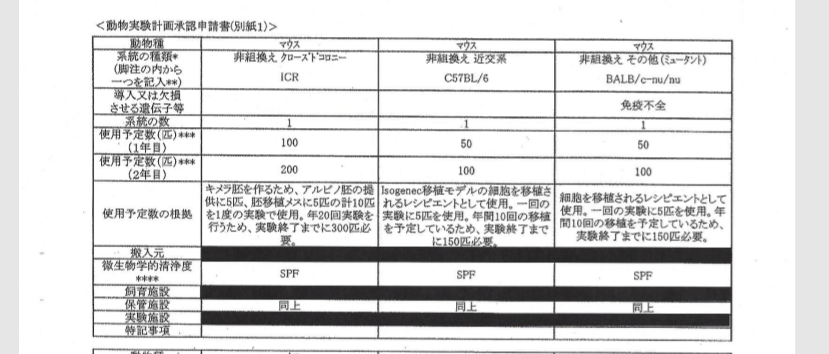
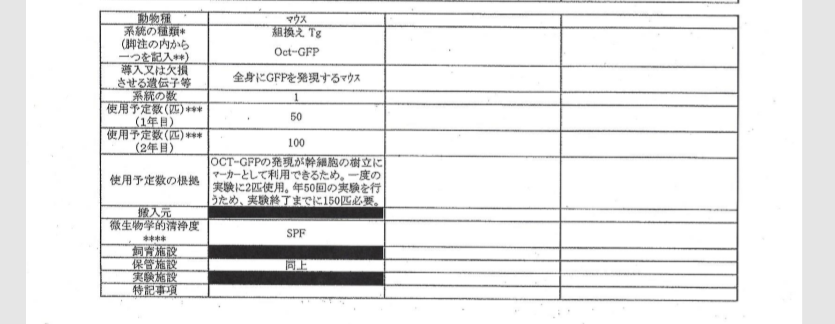
ICRはキメラのリシピエントマウス。B6とヌードマウスはテラトーマのリシピエントマウス。GOFマウスはSTAP細胞のドナーマウスで無論、若山研で自家飼育している分です。
和モガさんが書いているようにまだこの時はキメラ実験はしていませんからF1マウスの申請はしていない。あくまでもGOFマウスでキメラ実験する予定です。で、このことは手記にしか書かれていないことですが、GOFマウスでは失敗した。B6はESでも作りにくいマウスなのでF1でやってみようということになって、それで作ってみたら、又失敗した。
>>
2011年10月頃には、ATPに細胞を晒すことが試みた中では最も適したストレス条件であることもわかり、明るく光る(つまりOct4が強く発現している)細胞塊が観察されるようになっていた。顕微鏡下で見てもらうと、若山先生も「うん、ちゃんと光ってるね」と言ってくださった。こうして、「ちゃんと光る細胞塊」を確認してから、若山先生がキメラ作製の実験に力を入れてくださるようになった。(89P)
ここにまず時期の情報がある。キメラ実験の準備は2011年10月頃にちゃんと光る細胞塊が出来るようになった後である。和モガさんはこの手記の記載をしっかり確認していない。
①STAP動物実験計画書提出(2011.9.9)-Oct4-GFPマウスで申請しているので、この時は129B6F1マウスを使う予定はなかった。
②GOFマウスで実験-Oct4-GFPで最適な初期化方法を模索。-初期化方法が確立されたので、キメラが出来やすい129B6F1マウスに変更。
③129B6F1マウスで実験
④129B6F1マウスでキメラ・STAP幹細胞作製に成功。(2011.10月頃?)-マイクロナイフで切り、塊で入れて初めて成功。
ここまでの段階で既に日程が違っていますね。最初のキメラ成功は11月だと自己点検委員会の報告にもある。又、4Nキメラの写真の日付は2011/11/28です。10月頃に光ったというのは曖昧なので9月の後半まで含んでいてもいいでしょうが、光ってから準備を始めたと手記に書いてある。その時系列は以下の通りです。
>>
キメラマウス作製のためには卵の準備、スフェア細胞の準備、キメラ胚を移植するお母さんマウスの準備のすべてを、タイミングを合わせて行わねばならない。若山先生が計画表を作成してくださり、いよいよキメラマウス作製実験が本格的に始まった。しかし、何度試みていただいたものの、やはりES細胞から作製されるようなキメラマウスはできてこなかった。(90P)
計画表から作製したと書かれている。小保方さんはそれを見ている。そしてその計画表通りに実験は行われ、小保方さん自身も計画表の日程通りに酸浴細胞を作成したのです。小保方さん本人が確認したことを書いている。ここで何日かかったかが計算できる。
①計画表の作製は1日
②GOFマウスの赤ちゃんマウスを得るのに交配から始めて最低20日。
③酸浴細胞を作るのに最低7日。
④キメラ胚を得るのに交配から始めて4日。
⑤仮親の疑似妊娠状態を作るのに2日程度。
⑥キメラが生まれるまで移植後16日。
この6つの作業を同時並行に行って経過する最長時間は①③⑥の合計です。
約25日以上ですね。②は何時も準備されているから小保方さんはすぐ酸浴細胞の作製を始められるのです。そしてそれを受け取ってからでないと若山さんはキメラ胚インジェクションはできませんが、キメラ胚自体は酸浴細胞がインキュベーターの中にある7日間の間にジャストインタイムで作っておけばいい。仮親も同じです。インジェクトしたら1日休めてから仮親の子宮に入れる。そこから生まれるまで20日から4日を引いた16日です。
何度か試みたと書いてある。インジェクションは一日で終わってない。2,3日かけている。だからほぼ1か月かかっている。10月1日に始めたら11月1日に出来なかったという結果が出るのです。10月頃に光ったと書いているが、これが9月の20日だったら10月の20日ころに失敗確認ができたということになる。
和モガさんの以下の時系列認識はちがっていますよね。まだ成功してない。
>>
④129B6F1マウスでキメラ・STAP幹細胞作製に成功。(2011.10月頃?)-マイクロナイフで切り、塊で入れて初めて成功。
小保方さんの手記の続きを確認しましょう。
>>
私はもうES細胞からのようなキメラマウスはできないというのも重要な結果の一つと捉え、分化した体細胞がストレスを受けるとOct4陽性の細胞塊ができてくるまでの細胞の変化過程を論文化しようと考えていた。GOFのスフィア塊からはちゃんとしたキメラは出来なかった。博論時のキメラのようなGFP細胞が何個が死なずに残っているようなのではなくて、ES細胞からできるようなスタンダードなキメラは出来ないと確認したということです。これが早ければ10/20頃、遅ければ10月の末か1日ころだということになる。
ところがここからF1マウスでの実験が始まったのです。手記の続きを確認しましょう。
>>
ところが若山先生はどうにかスフェアからキメラマウスを作製する方法を思案してくださっていた。イヌに多種の血統があるようにマウスにも多種の系統があり、実験結果が安定するように各系統のマウスの系統は厳密に管理されている。ここは近交系マウスの説明を若山さんから受けたんでしょうね。F1でやってみることになる前振りです。小保方さんはGOFでできなかっただけであきらめたのですから、そういうことは知らなかったでしょう。手記は続きます。
>>
ある日、若山先生から、「ES細胞にはES細胞が樹立しやすい系統のマウスと樹立が難しい系統のマウスが存在している。また、クローンマウスを作製するときにも、クローンマウスを作りやすい系統のマウスが存在しているから、その系統のマウスを使って小保方さんのキメラマウスの作製を行ってみたい」とご提案いただいた。ここの記載は注意深く読んでいただきたいが、若山さんはクローンマウスを作りやすい系統のマウスを使って小保方さんのキメラマウスを作成したいと言っている。小保方さんの細胞でクローンマウスを作るのだとも聞こえる言い方ですね。
何か、小保方さんを教育でもしているような言い方にもなっている。気になるところです。彼は小保方さんを一度リクルートしようとしてヴァカンティに反対されている。本当なら小保方さんを受け入れなければいいだけですが、その後も引き続きGOFマウスを使わせるという名目で彼女を客員受け入れしている。小島さんまで名目上の客員にしている。
私の説だと若山さんはここで小保方核使用ntES実験に切り替えたということになるんですが、今は和モガさんの仮説検証ですから、ここでは言わない。続きです。
>>
若山先生が準備してくれたマウスは129xB6 F1と呼ばれるマウスで、129という系統のマウスとB6という系統のマウスを交配させて作製した雑種の赤ちゃんマウスだった。
今回のキメラ実験は、まず雑種の赤ちゃんマウスの準備、そのマウスを使ったスフェア作製の日程の決定、卵の準備、移植するお母さんの準備をすべて若山先生が計画し準備してくださった。キメラ実験を行う1週間前に若山先生から赤ちゃんマウスを渡され、スフィアを作成してキメラ実験用に若山先生に渡すように指示を受けた。何度か繰り返し実験を行ったが、やはりキメラマウスはできてこなかった。それにもかかわらず、若山先生はあきらめずに実験を繰り返してくださった。どうでしょうか。キメラマウスが出来てこなかったと確認できたのは計算上何時頃になるでしょうかね。計画に1日、F1マウスの交配から始めて赤ちゃんマウスを得るのに最低でも20日かかる。そこからスフィア作成に7日かけて移植に1日、そして1日休めて、子宮に移す。そこから最低でも16日で出産。仮に10日胚で帝王切開するとしても、6日かかる。計35日です。前回のGOFでの失敗確認が早くて10月20日ころでした。11月25日ころに帝王切開して失敗確認したということになる。さて、続きです。いよいよ成功しますね。
>>
ある日いつもの通りスフェアを渡すと、「これまではスフェアをバラバラの細胞にしてから初期胚に注入していたが、今日からはマイクロナイフで切って小さくした細胞塊を初期胚に注入してキメラマウスを作ることにした」とおっしゃった。それから10日後、若山先生からキメラができたと連絡を受けた。その上。残りの細胞をES細胞樹立用の培養液で培養したらES細胞様に増えだしたと報告された。毎日、スフェア細胞を培養し観察していた私は、細胞が増える気配すら感じたことがなかったので大変驚いた。「特殊な手技を使って作製しているから、僕がいなければなかなか再現がとれないよ。世界はなかなか追いついてこられないはず」と若山先生は笑顔で話していた。私の説では若山さんはいたずらしているので笑顔だったと自然に理解してますけどね。まあ、ここでは和モガ説批判ですから控えて置きましょう。
F1での最初の実験結果が失敗だと確認できたのは11月25日前後だという計算になっていましたね。でも記録に残っている最初のキメラは11月28日の4Nキメラです。10日の胎児だとして、ここから小保方さんの説明している経緯を逆算して行けます。
11月28日 10日胚の帝王切開、蛍光顕微鏡撮影
11月27日 9日胚
11月26日 8日胚
11月25日 7日胚
11月24日 6日胚
11月23日 5日胚
11月22日 4日胚 仮親子宮への挿入
11月21日 4日胚 一日培養
11月20日 4日胚 リシピエント胚盤胞へのスフィア細胞移植(今日からナイフ切り分け)
11月19日 7日目 インキュベーター内
11月18日 6日目 インキュベーター内
11月17日 5日目 インキュベーター内
11月16日 4日目 インキュベーター内
11月15日 3日目 インキュベーター内
11月14日 2日目 インキュベーター内
11月13日 1日目 F1マウスATP酸浴
記録に残っている最初のキメラの赤ちゃんマウスを小保方さんが受け取ったのは11月13日です。そしてこれは成功したキメラですから、失敗しているキメラ実験はこれより前の段階で渡されている。小保方さんはF1でやることになってからはずっと赤ちゃんマウスを渡されつづけていて毎日スフィア酸浴をしているからこの時期はずっと若山さんに細胞を提供し続けることができる。
小保方さんは以下のように書いている。
>>
何度か繰り返し実験を行ったが、やはりキメラマウスはできてこなかった。それにもかかわらず、若山先生はあきらめずに実験を繰り返してくださった。キメラが出来てこないがあきらめずに実験を繰り返していたが11月20日にはナイフ切り分けでやると言った。つまり以前の結果が出来てないからです。前日作っているのはまだ結果が出てないかもしれないが、前日までに結果が分かったものに関しては出来てなかったから、11月20日にナイフ切り分けと決めた。最後に出来てないと判断したキメラに移植したスフィア細胞は11月12日に小保方さんが酸浴させたものということになる。するとこれより数日前までは失敗続きだったのだから、11月5日頃から小保方さんはF1マウスの赤ちゃんを受け取っていたことになる。
もう一度日程を確認してみよう。今度は逆算でなく10月1日を起点にしてみる。
10月1日 GOFキメラ作製計画書作製。
10月2日 GOF赤ちゃんマウスATP酸浴
10月3日 インキュベーター1日目
10月4日 インキュベーター2日目
10月5日 インキュベーター3日目
10月6日 インキュベーター4日目
10月7日 インキュベーター5日目
10月8日 インキュベーター6日目
10月9日 インキュベーター7日目
10月10日 リシピエント胚盤胞胚へスフィア細胞移植
10月11日 リシピエント胚盤胞胚一日培養
10月12日 リシピエント胚盤胞胚を仮親子宮に挿入
10月13日 キメラ5日胚
10月14日 キメラ6日胚
10月15日 キメラ7日胚
10月16日 キメラ8日胚
10月17日 キメラ9日胚
10月18日 キメラ10日胚
10月19日 キメラ11日胚
10月20日 キメラ12日胚
10月21日 キメラ13日胚
10月22日 キメラ14日胚
10月23日 キメラ15日胚
10月24日 キメラ16日胚
10月25日 キメラ17日胚
10月26日 キメラ18日胚
10月27日 キメラ19日胚
10月28日 キメラ20日胚自然分娩
10月29日 1日遅れキメラの自然分娩
10月30日 2日遅れキメラの自然分娩
10月30日 3日遅れキメラの自然分娩
10月31日 4日遅れキメラの自然分娩
11月1日 5日遅れキメラの自然分娩
11月2日 F1マウスキメラ作製計画書
11月3日 F1マウス交配
11月4日 F1マウスプラグ確認(妊娠確認)
11月5日 F1マウス妊娠2日目
11月6日 F1マウス妊娠3日目
11月7日 F1マウス妊娠4日目
11月8日 F1マウス妊娠5日目
11月9日 F1マウス妊娠6日目
11月10日 F1マウス妊娠7日目
11月11日 F1マウス妊娠8日目
11月12日 F1マウス妊娠9日目
11月13日 F1マウス妊娠10日目
11月14日 F1マウス妊娠11日目
11月15日 F1マウス妊娠12日目
11月16日 F1マウス妊娠13日目
11月17日 F1マウス妊娠14日目
11月18日 F1マウス妊娠15日目
11月19日 F1マウス妊娠16日目
11月20日 F1マウス妊娠17日目
11月21日 F1マウス妊娠18日目
11月22日 F1マウス妊娠19日目
11月23日 F1マウス妊娠20日目出産11月24日 F1赤ちゃんマウスATP酸浴
11月25日 インキュベーター内2日目
11月26日 インキュベーター内3日目
11月27日 インキュベーター内4日目
11月28日 インキュベーター内5日目
11月29日 インキュベーター内6日目
11月30日 インキュベーター内7日目
12月1日 リシピエント胚盤胞胚へスフィア細胞移植
12月2日 リシピエント胚盤胞胚一日培養
12月3日 リシピエント胚盤胞胚を仮親子宮に挿入
12月4日 4Nキメラ5日胚
12月5日 4Nキメラ6日胚
12月6日 4Nキメラ7日胚
12月7日 4Nキメラ8日胚
12月8日 4Nキメラ9日胚
12月9日 4Nキメラ10日胚 帝王切開胎児確認
12月9日 1日遅れ4Nキメラの帝王切開確認
12月9日 2日遅れ4Nキメラの帝王切開確認
12月9日 3日遅れ4Nキメラの帝王切開確認
12月9日 4日遅れ4Nキメラの帝王切開確認
12月9日 5日遅れ4Nキメラの帝王切開確認
12月10日 今日からナイフ切り分け、リシピエント胚盤胞胚へスフィア細胞移植
12月11日 リシピエント胚盤胞胚一日培養
12月12日 リシピエント胚盤胞胚を仮親子宮に挿入
12月13日 4Nキメラ5日胚
12月14日 4Nキメラ6日胚
12月15日 4Nキメラ7日胚
12月16日 4Nキメラ8日胚
12月17日 4Nキメラ9日胚
12月18日 4Nキメラ10日胚 帝王切開胎児確認成功(記録では2011/11/27)
つまり記録されているキメラのできた2011/11/28日付が最初のキメラだとすると21日合わないことになるので、スタートの10月1日仮定を9月9日に修正すればいいことになる。小保方さんが10月頃に良く光るようになったと書いているのを9月の前半だったと解釈する。
もしくは、一番自然なのはF1マウスを1から作ったかということを疑えばF1マウスは既に用意されていたのだと考えて、小保方さんが疑っていないから、早すぎると気付かなかったと考える。最大20日縮まることになる。これだとぴったり合うことになる。
(和モガ第三者説批判)ここまで日程を押さえた上で和モガさんのフローチャートを検証してみよう。
最初の段階で既に無理があると分かる。2011.10月頃という記述が既に最初のキメラは11月だという理研の発表と整合していない。レターの写真が2011/11/28撮影のキメラだという理研の説明も考慮されてない。
ただし、あくまでも?マークなので、既述した日程で小保方さんの勘違いで9月9日に良く光って、かつF1は既に準備されていたと考えると21日と20日で41日早まる。すると12月18日から41日遡ると、11月7日になる。ここで11/28のキメラ以前に最初のキメラが出来ていたのだと考えることもできる。桂報告書と若山さんは11/28のキメラが最初のキメラとは言ってない。ただ11月に最初のキメラが出来たと言ってるだけなので、11/7から11/28までの間に最初のキメラがあった可能性はある。
さて、やっとOoboe さんのご要望に応えて、和モガ説批判を述べることが出来る準備が整った。2011.10月頃?は11/7~11/28と読み替えましょう。もう一度確認します。
①STAP動物実験計画書提出(2011.9.9)-Oct4-GFPマウスで申請しているので、この時は129B6F1マウスを使う予定はなかった。
②GOFマウスで実験-Oct4-GFPで最適な初期化方法を模索。-初期化方法が確立されたので、キメラが出来やすい129B6F1マウスに変更。
③129B6F1マウスで実験
④129B6F1マウスでキメラ・STAP幹細胞作製に成功。(2011.10月頃?)-マイクロナイフで切り、塊で入れて初めて成功。
この後に第三者が捏造するという説なのです。
⑤STAP細胞の成功に嫉妬した犯人が、この後、STAP論文を潰す行動に出る。
成功に嫉妬するというからには成功を知っている人ですからまずはラボメンバーです。小さいラボです。常勤メンバーは若山さんと小保方さんは成功に嫉妬する第三者ではあり得ませんから、奥さん、野老さん、李さん、学生の京極さん、学生の糸井さん、女子学生の寺下さん、テクニカルスタッフの坂出さん、同じく、山中さん、事務の平内さんだけです。
奥さんが御主人の成功に嫉妬するでしょうかね。事務の平内さんはそうい仕事にはタッチできません。糸井さんは小保方さんにGOFのntESを一皿くれた人です。2012年には大学に戻っています。京極さんは最後まで小保方細胞で自分の論文を書くように若山さんから指導されていた人だと手記にある。FLS-T1,2の作成時に若山さんと一緒に小保方さんから作り方を習った人です。野老さんと寺下さんはレター論文の共著者です。
残るのは李さんとテクニカルスタッフの二人の女性だけですが、坂出さんは大田さんとの共著論文もある位のベテランです。李さんはFLS-T1,2の実験時に日本に居ません。山中さんはずっといましたけどね。
でも、第三者ってそもそもあるのでしょうかね。次に和モガさんはその手口について書いている。
⑥若山B6マウスを岡部B6マウス(Acr/CAG-GFP)にすり替え。
成功に嫉妬したために若山B6マウス(CAG-GFP)を岡部B6マウス(Acr/CAG-GFP)にすり替えたというが、若山さんは自分の記者会見でGOFマウスはテクニカルスタッフに任せることもあるが、他のマウスは自分が管理していると答えていて、交配も自分で行うと証言している。和モガさんの説ではこれは第三者を庇う嘘だということになる。若山さんにとってこういう悪事をする人を庇わなければならないとしたら奥さんしかないでしょう。他は単なるいわば使用人です。首にすれば済む話だ。でも、では奥さんがどうしてご主人の関与して成功している実験結果に嫉妬するのでしょうかね。
⑥若山B6マウスを岡部B6マウス(Acr/CAG-GFP)にすり替え。
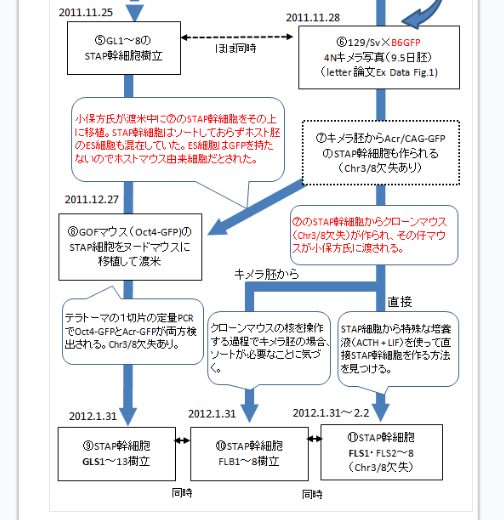
の後に
⑥若山B6マウスを岡部B6マウス(Acr/CAG-GFP)にすり替え。129xB6GFP 4Nキメラ写真(9.5日胚) (letter 論文 Ex Data Fig 1)
が出来たと。こうやって見てくると和モガさん最初の記述より修正してきているね。でもちゃんと論理が整合していればいい。
ただ、これでも実験を妨害するためにB6を入れ替えたとする推測には4つの難点が残っている。
1.これで妨害したことになってるか。つまり、これが発覚したのは3年後であるが。
2.F1は若山さんしか管理できないと若山さん自身が言ってる。
3.庇われている筈の奥さんには動機がない。成功は生計を一にしている者にとっては喜ばしい。
4.李さんと山中さんがつるんでいたとして、2011/11/28の実験、翌年のFLS、FLB、GLS、胎盤が光ったときの2012年春のキメラ、そしてAC129、更にFLS-T1,2の全てに若山さんの目を逃れて偽マウス、或は「僕のマウス」ESを渡せるか。
続けてフローチャートを下って行こう。
⑦キメラ胚からAcr/CAG-GFPのSTAP幹細胞も作られる。(Chr3/8欠失あり)
⑧2011312.27 GOFマウス(Oct4-GFP)のSTAP細胞をヌードマウスに移植して渡米 小保方氏が渡米中に⑦のSTAP幹細胞をその上に移植。STAP幹細胞はソートしておらずホスト胚のES細胞も混在していた。ES細胞はGFPを持たないのでホストマウス由来細胞だとされた。
⑦は⑥と同時に作られていて増殖確認をするだけだから3継代もすると一応増殖しているという判断はできる。正式な確認は後でちゃんとやればいい。後に小保方さんの行ったFLS1-8全ラインの40継代がそれである。因みに試料が残されているのに、肝心なFLSの実験を小保方さんが行っていることは桂報告は触れずに、ES細胞のグラフを2011年の春に行ったという小保方さんの嘘だけを追求した。震災避難で腰かけている時にES細胞の増殖確認なんかするわけもなければ、そんな分かり切った実験に若山さんが予算を使わせるわけもないことである。
キメラが出来たことを確認できたときには増殖実験も確認できているので、⑥⑦は同時である。
(Chr3/8欠失あり)というのはB6の3番と129の8番にある欠失のことであるが、言うまでもなく、この幹細胞は残されていないのであるから、⑦に書き込むのは誤解を生むかもしれない。ただ⑧のテラトーマからは(Chr3/8欠失あり)が検証されているから、犯人が小保方さんのテラトーマの上から⑦の幹細胞を注射したのなら、⑦の幹細胞には(Chr3/8欠失あり)だったのだろうという演繹推論である。他にテラトーマに入れられた(Chr3/8欠失あり)の細胞があるならば、この時の幹細胞が(Chr3/8欠失あり)だったかどうかは分からないことになる。その証拠に桂報告書は大田ESを小保方さんが注射したと言ってる。どちらが正しいのか。
小保方さんは学生のGOF-ESを持っているので捏造するにしても太田ESを注射するなんて馬鹿げ過ぎている。桂チームと言うのはアルツハイマー集団だと思われる。
まあ、従って、私もここは最初の幹細胞が注射されたと思っているが、私は若山さんが注射した、和モガさんは第三者が注射したと主張しているのである。若山さんの作った筈のこの最初の大事な幹細胞を誰が失敬してテラトーマの各箇所に注射できるのか。私には若山さん本人しか思いつけない。
ここで幹細胞の作り方に関して和モガさんはキメラ胚を使ったと考えて居る。ナイフ切り分けでたくさんの胚盤胞移植が行われ、そのうちの一部は仮親の子宮に挿入されてキメラとなったが、余ったSTAP塊移植後の胚盤胞はしばらく培養された後インナーセルマスを取り出されて、ES培地で培養されたと考えて居る。
インナーセルマスはリシピエントのそもそも持っているインナーセルマスと移植されたSTAP細胞が混在している。所謂共培養状態になっている。
これを取り出してGFPでFACS選別したのが完成されたSTAP幹細胞だという説である。これはクムリナ書き込みから推測されている。
私はクムリナ書き込みはカモフラージュだとしか思ってない。彼は小保方核使用ntESをつくって、その成否をキメラ胚に入れて確認しただけである。キメラが出来ればその株はntES化に成功しているということになる。ntESは培養できただけでは駄目でキメラ樹立まで確認されないといけない。ここは私もなぜキメラ胚に一度入れるのかが分かってなかった時期がある。なんだか自然に分かってきた。
さてここまででキメラと幹細胞はできた。和モガ説であると最初のキメラは若山さんがナイフ切り分けで作ったのである。そしてこの時の幹細胞も若山さんが小保方さんに出来たといったのだからどういう作り方はともかくとしてできた。本当の成功である。
その後に、第三者が若山B6を岡部B6に取り換えた。(F1を交配したのは若山さんで、GOF以外はテクニカルスタッフに任せてないと記者会見で答えているのだが、第三者は奥さんということなのであろうか。奥さんが旦那の成功を妬んだと?)
ということは最初の成功マウスは若B6-CAGだったという推定であるが、誰かがそんなことを言ったであろうか。若山さんはFLSに関して「僕のマウス」を渡したと証言しただけで、前年度の最初の実験でどんなマウスを使ったか語ってない。ただ、2011/11/28撮影の4Nキメラに関して129/Sv x B6GFPだと答えただけである。
僕のマウスというのは129-CAG-GFP x B6-CAG-GFPのF1マウスのことである。
第三者は最初の実験で若山さんが129 x B6-CAG-GFPのF1マウスを使って成功したのだと知っていることになる。そして成功を羨んで妨害しようとして次の実験の時に129 x B6-Acr-CAG-GFPのF1マウスを渡したという。妨害者は三年後に理研の調査でアクロシンが発見できるように妨害しようとしたというのか。途中で小保方さんが理研に拾われなかったら、アクロシンマウスは発見されなかったかも知れない。何を妨害したことになるのであろうか。そもそも妨害されなくても三誌リジェクトされている。
フローチャートを下ると以下のように続く。
⑦キメラ胚からAcr/CAG-GFPのSTAP幹細胞も作られる。(Chr3/8欠失あり)
↓
⑦のSTAP幹細胞からクローンマウス(Chr3/8欠失)が作られ、その仔マウスが小保方氏に渡される。
誰がクローンマウスを作ったのか? 第三者がクローンマウスを作って小保方さんに渡したのか。仔マウスは若山さんが交配して後、彼自身が小保方さんに渡すと記者会見で述べられている。
2011/11/28の幹細胞からクローンマウスを作るのに20日かかる。2011/12/18である。その仔を作るには親の成熟を待たねばならない。早くても50日かかる。そこから交配して更に20日後に仔マウスが得られる。
ここに無茶な推定が入っているのだけれども、逆に、なぜ和モガさんはこういうクローンを考えなければならないのかと推測すると、翌年の初頭の実験に関して若山さんが「僕のマウス」を渡したと言ってるから、若山さんは騙されて実は第三者がアクロシン入りマウスに取り換えたとしなければならないからであろう。そもそもが最初から使われているB6はアクロシン入りで、若山さんが嘘をついて「僕のマウス」を渡したのだと言ってるというストーリーでないところから来る無理ではないかな。
私はクローン胚からntES化していると考えて居る。これだと無理がない。犯人は若山さんです。
それを若山さんは無実で最初成功していたのに、第三者が嫉妬して、アクロシンマウスと取り換えたというストーリーに対して、事実関係の情報が不整合になるのだと思われる。そもそもアクロシンマウスに取り換えることが当面の妨害にすらなってないではないかと疑義されるところでもある。アクロシンが問題になったのは3年後ですよ。しかも、あのまま小保方さんが渡米したままだったら何の問題も起きてない。
取り敢えず、Ooboe さんのご依頼に対する回答はこのくらいにしておきましょう。Ts.Marker さんには別途コメント欄に書いて置きましょう。
①桂報告書---キメラと幹細胞の既存ESコンタミ捏造説は完全論破されている。
②第三者妨害説---和モガ説。今批判した。
③キメラは論文通りにできている。幹細胞は論文通りではなく共培養で作られた。---Ts.Marker 説
④キメラも幹細胞も論文通りにできている。---Ooboeさんとパートナー氏説。検察申請中。
⑤キメラと幹細胞は小保方核使用ntES---居士説。
- 2020/01/25(土) 09:23:25|
- ミトコンドリア
-
-
| コメント:9
Ts.Marker 氏がSTAP関連細胞株RNA公開データベース情報内に存在していたmtDNAの一変異の存在に関して報告されている。AC129に関する問題とは別にここで検討したい。
この事実が報告されたのは2017/11/23である。以下、彼のブログである。>>
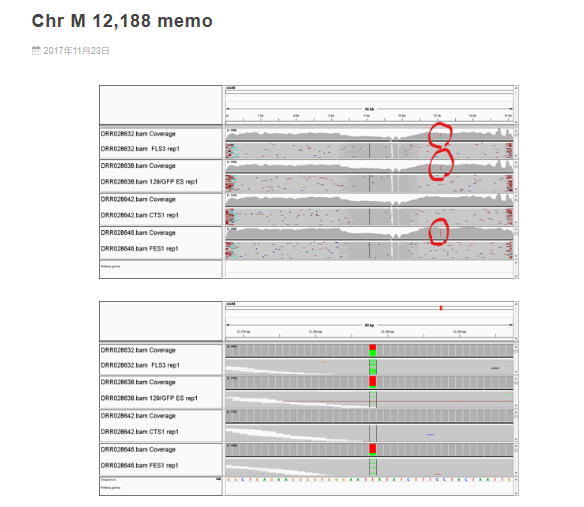
たった16Kbしかないミトコンドリアでくっきり1箇所。
CTS1だけない。 確定かな?
Tru-seq
SRR1171560 ESC T:1572(100%) A:0
SRR1171580 STAP T:2396(100%) A:1
SRR1171585 STAP-SC(FLS?Acr) T:1245(85%) A:217(15%) ??
SRR1171565 FI-SC(B6+α?) T:2212(100%) A:2
SRR1171590 TSC T:2359(100%) A:0
SRR1171556 CD45+ T:2000(100%) A:1
SRR1171558 Epi-SC T:1103(100%) A:0
ChIP-seq input
SRR1171564 ESC T: 191(100%) A:0
SRR1171584 STAP T: 178(100%) A:0
SRR1171589 STAP-SC T: 171(100%) A:0
SRR1171569 FI-SC(CTS?Acr) T: 144(100%) A:0
SRR1171594 TSC T: 229(100%) A:0
SMARTer-seq
SRR1171574 ESC T:1396(100%) A:0
SRR1171578 STAP T: 970(100%) A:1
WGS
DRR028646 FES1 T:3625(80%) A:921(20%)
DRR028638 129/GFP ES T:2963(80%) A:733(20%)
DRR028632 FLS3 T:1949(49%) A:1992(50%)
DRR028633 FLS3 T:1912(49%) A:2015(51%)
DRR028642 CTS1 T:3718(100%) A:4
DRR028643 CTS1 T:3667(100%) A:5
WGSの残りはマッピング中。時間がかかる.....
------------- 11/26 追加 -----------------------
DRR028652 FES2 T:5307(100%) A:4
DRR028656 129CAG T:1219(100%) A:0
(Low coverage sequencing of 129 and C57BL/6)
WGS Illumina HiSeq 2000- Peter Sorger-Harvard Medical School
SRR1614029 129X1/SvJ T:3(100%) A:0 G:0 C:0
------------- 11/29 追加 -----------------------
DRR028644 ntESG1 T:4990(100%) A:4
-------------- 12/5 追加 -----------------------
DRR028650 ntESG2 T:4291(100%) A:1
SRR2917581 ICR ? T:219(100%) A:0
-------------- 12/7 調べ -----------------------
Mouse strain assembly hub - May 3, 2017 より
DBA/2J T
129S1/SvImJ T
BALB/cJ T
C57BL/6NJ T
NOD/ShiLtJ A
 「確定かな?」は何も説明が無く、独り言の類なので何も人には伝わらない。ただ、ここには一個の事実が提示されているということである。
「確定かな?」は何も説明が無く、独り言の類なので何も人には伝わらない。ただ、ここには一個の事実が提示されているということである。
この事実を自分なりに理解しようと努力された最初の人が和モガ氏であった。以下、和モガ氏のブログである。>>
「
STAP細胞事件」-FLS3にはクローンの痕跡がある
2017/11/20(月) 午前 9:02にTs.Maker氏のブログのコメントに次のコメントが書かれていた。Ts.Maker氏自身のコメントである。
「memo chrM 12,188」
「chrM」とはミトコンドリアのDNAを指し、何かを見つけたのでその地番を12,188とメモ書きしたということであろう。
体細胞は核DNAの他にミトコンドリアに独自のDNA(mtDNA)を持つ。このmtDNAは必ず母親から子に受け継がれ、父親から受け継がれることはない。したがってmtDNAを調べれば、母親、母親の母親、さらに母の母の母と女系を遡ることができる。またmtDNAは組換えが起こらないので、mtDNAに違いがあるとすればそれは突然変異によるということになる。
私はFLS3がクローンマウスの仔から作られていると考えていたので、FLS3がクローンならミトコンドリアは移植先のmtDNA(おそらくICRマウス)になり、ミトコンドリアを調べれば129B6F1の母親側の129マウスとは違う塩基(SNP)が見つかるだろうと思っていた。そこで、このコメントを見て、すぐFLS3のミトコンドリアを見にいった。
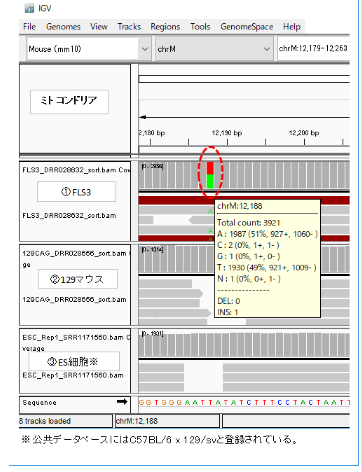
IGVで見ると①のようになっていた。これは予想外であった。①のFLS3に塩基AとTが半々ずつ出ていたからである。mtDNAは母親のDNAしか持っていないので、1塩基中に2種類出るとは思っていなかったのである。
しかし、文献「体細胞クローン金華豚、およびその後代産子におけるドナー体細胞由来mtDNAの伝達性」を見ると、次の記述があり、体細胞クローンではドナーとレシピエントのmtDNAが混在すると書かれていた。
体細胞クローンウシでドナー由来のmtDNAが検出されて(Hiendlederら1999;Steinbornら2000)以来、体細胞クローン動物に関しては、伝達割合に差はあるものの、クローン個体内でドナーとレシピエント双方由来のmtDNAが混在する状態(ヘテロプラズミー)と考えられつつある(Takedaら」2000;Inoueら2004)
AとTが両方でるのは、FLS3が体細胞クローンで12,188番のTの塩基はドナーの129B6F1の129系塩基T(②)に、レシピエントのmtDNA(おそらくICRマウスで塩基はA)が混在しているからだと思われる。
調査委員会はFLS3はFES1由来で、FES1は129×B6の受精卵ES細胞ということであった。それなら、mtDNAの12,188は②のように塩基Tで灰色表示されるはずである。作製者の太田氏は作成後、研究には使わなかったというので、突然変異が半分まで蓄積するはずもない。FLS3が受精卵ES細胞でないのは明らかだ。
もし、FES1のミトコンドリアもFLS3と同じなら、桂調査委員会が調べたFES1は太田氏の作成した受精卵ES細胞ではなく、すり替えられ太田氏作製のFES1として調査委員会に提出されていたことになる。逆に、FES1がFLS3と違うなら、それはそれで、FLS3はFES1由来とは言えないことになる。
いずれにしてもこのミトコンドリアの一塩基で桂調査委員会は詰んでいるのである。
2017.11.21 Tue l STAP細胞事件 l コメント (1) トラックバック (0) l topICRがAであろうという予想は上のTs.Marker氏のデータ上では外れている。以下、その2である。>>
「STAP細胞事件」-ミトコンドリアの特異点はクローンマウスの痕跡である(2)
この129B6F1 Acr/GAG-GFPの細胞株だけを抜き出すと次のようになる。
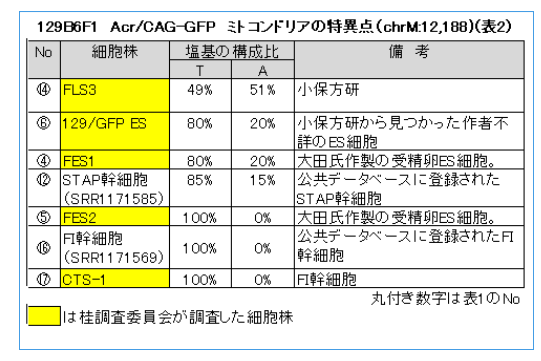
それぞれの細胞株は塩基のT(チミン)が49%から100%まであり、それに伴いA(アデニン)が51%から0%になっている。
実は、「FLS3にはクローンの痕跡がある」を書いてから、もしかしたらFES1の特異点に二重塩基はないのではないかと思っていた。それは、文献「ミトコンドリア DNA 複製の常識の一端が覆る」に次の記述を見つけたからである。
個々の細胞には、数百から数千個以上ものミトコンドリアゲノムDNAがあり、しばしば突然変異することから、このDNAの組成はかなり不均一になると予測されます。また、歳をとると、一部の組織のミトコンドリアDNAは不均一になっていきます。しかし奇妙なことに、増殖している個々の細胞や新生児では全身で、全てのミトコンドリアDNAが同一の遺伝子型組成(同一のDNA配列)を持っています。この「ホモプラスミー」と呼ばれる状態へ短時間にリセットするという細胞質遺伝の現象は、酵母からヒトまで共通にみられます。
クローンの仔はドナーとレシピエント両方のミトコンドリアDNAが混在する状態(ヘテロプラズミー)で生まれてくる。このため、クローンマウスから作ったSTAP細胞のミトコンドリアはヘテロプラズミーである。そこからSTAP細胞を培養してSTAP幹細胞を作ると、その培養は細胞の形質を変える培養なのでミトコンドリアはそのままであろう。
しかし、いったん出来たSTAP幹細胞を培養するとSTAP幹細胞は増殖するので、ミトコンドリアがホモプラスミー化することになる。
つまり、クローンマウスからFLS3が作られヘテロプラズミーとなったミトコンドリアはFLS3→129/GFP ES→FES1と培養していくにつれ、次第にホモプラスミー化していき、FES1では特異点の二重塩基がみられない可能性もあると思っていたのだ。
実際には、④FES1と⑥129/GFP ESはT:80%、A:20%で共に特異点の二重塩基が検出されてる。公共データベースに登録されていた⑫STAP幹細胞はT:85%、A:15%で、Aの構成比がそれより低い。これらは、培養によるミトコンドリアのホモプラスミー化で説明できるのである。
⑤FES2については市販の129X1雌マウスとAcr/CAG-GFP B6雄マウス(岡部マウス)から作られた受精卵ES細胞だろうと思う。このため、二重塩基にはならなかった。
一方、⑯FI幹細胞、⑰CTS-1の3つの細胞株では特異点の二重塩基が消えているが、これについては完全にホモプラズミー化したということではなく、別の理由だと思う。
ついては完全にホモプラズミー化したということではなく、別の理由だと思う。
2018.01.21 Sun l STAP細胞事件 l コメント (0) トラックバック (0) l topその3である。>>
「STAP細胞事件」-ミトコンドリアの特異点はクローンマウスの痕跡である(3)
以下、「ミトコンドリアの特異点はクローンマウスの痕跡である(1)」で書いた「②特異点が塩基『A』を持つマウスの系統」の話である。
DNAにはその塩基の種類を見れば、マウスの系統が分かるSNPs(一塩基多型)が存在する。
FLS3が129X1雌マウスとB6雄マウスの受精卵ES細胞なら、ミトコンドリアDNAは雌の129X1になっているはずで、どこかに129X1マウス系統のSNPsを検出できるはずである。
ところが、ミトコンドリアDNAには129X1と分かるSNPsが存在しない。ミトコンドリアのDNAはわずか16,500塩基ほどで、常染色体の約一万分の一と桁違いに少ないのである。
SNPデータベースでは129X1を含めマウス系統の約90種類のSNPsが調べられるが、ミトコンドリアの特異点(12,188)が塩基「A」になるものはない。そもそも特異点はSNPsとは認識されていないのである。
その中で塩基「A」になっているマウスを調べ出したTs.Marker氏の情報は貴重なものであった。それによると、マウス系統は「NOD / ShiLtJ」である。もっとも、NOD / ShiLtJもSNPデータベースに登録されているマウス系統のひとつであるが、12,188にSNPsはないのである。

特異点(12,188)が塩基「A」になっている
特異点が「A」のマウスが存在するというのは非常に重要だ。このようなマウスがホスト胚であれば、クローンマウスはFLS3のようにヘテロプラズミーで生まれてくることになる。
NOD / ShiLtJマウスを提供しているCharles River社のホームページにはNOD / ShiLtJマウスは非近交系のICRマウスが起源であると書かれている。
実験マウスはクローズドコロニーと近交系の 2 種類に大別される。近交系は兄妹交配を 20 世代以上継続した系統で、系統内の個体はまったく同じ遺伝子組成をもつ。このため、SNPデータベースにSNPsを登録することが可能である。
一方、クローズドコロニーは 5 年以上外部から種マウスを導入することなく、一定の集団内でランダム交配により維持されている系統で、このため、各個体の遺伝的性質はばらつきがある。従って、マウス系統としてのSNPsを登録できないことになる。
若山研のクローン研究に伴う「動物実験計画承認申請書」ではクローズドコロニーマウスのICRマウスが申請されており、その数は年間、2,000個である。これだけの量ならどのタイミングでもクローンをつくることは可能だろう。
1月31日から2月2日にかけて作ったFLS3のクローンマウスのホスト胚は、たまたま特異点が「A」のICRマウスであった。その後、5月25日、7月9日のCTS-1、11~13も同じICRマウスが使われたが、このときのマウスの特異点は「A」ではなく一般的な「T」でヘテロプラズミーにはならなかったのだろうと思う。
2018.01.29 Mon l STAP細胞事件 l コメント (0) トラックバック (0) l top最近のTs.Marker 氏の書き込みである。>>
heteroplasmy の違いから
2020年01月04日
前記事 Chr M 12,188 memo(http://zscan4.livedoor.blog/archives/4664712.html)
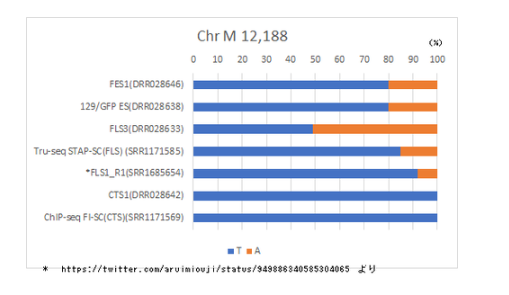
登録されたWGSデータから得られた.bamよりミトコンドリア(ChrM)のPosition12,188には、
通常 Tのところに変異したAが混在するヘテロプラスミーが見られる。
DRR028646 FES1 T:3625(80%) A:921(20%)
DRR028638 129/GFP ES T:2963(80%) A:733(20%)
DRR028632 FLS3 T:1949(49%) A:1992(50%)
DRR028642 CTS1 T:3718(100%) A:4
同じメスからの受精卵でもその比率はバラバラになるという。FES1と129/GFP ES 少なくとも同じヘテロプラスミーをもつ129のメスが起源。
比率がここまで同じだとやっぱりFES1の株分けかな。誰が持ってたかはわからないが...。
母系遺伝のヘテロプラスミーの比率を変化させるには
1.卵子形成or発生の段階で初期化
2.軽度の酸化ストレス
3.過酸化水素添加
4.核移植
など。
CTS1でAがほぼゼロになっているので核移植が疑われるが、FLS3でAの割合が逆に増えることの説明がつかない。
FLS3ではAの比率が上がりCTS1ではほぼゼロと傾向がバラバラになる可能性の一つは、STAP処理。
" ...the stress comprises removing at least about 40% of the mitochondria from the cell... "
と特許にあるようにSTAP処理では細胞質・ミトコンドリアをかなりの量を細胞から流出させる。
ミトコンドリアを半減近くまで減らせば、細胞ごとのヘテロプラスミーの比率はバラバラになるんじゃないかな。
ちなみにFLSの何番かはわからないが、Tru-seqのFLSと思われるデータは
SRR1171585 STAP-SC(FLS?Acr) T:1245(85%) A:217(15%)
とFES1や129/GFP ESに近いように見えるが5%は結構差がある。
つづく...。
つづき..
また、3の過酸化水素による理研の特許の実施例では「MELAS細胞に、活性酸素種として過酸化水素を接触させた場合、ヘテロプラスミー状態にどのような影響が生じるか検討した。 MELAS細胞(細胞密度;1×105〜1×106cell/mL)の培地中に最終濃度100μMとなるように過酸化水素を添加し、37℃、5%CO2の条件下で30分間インキュベートした後、細胞をPBSで3回洗浄し、3日間培養した。3日間の培養後、細胞密度細胞密度が5×105〜6×106cells/mLに達した後、104〜1.2×105倍に希釈してプレーティングを行い、さらに、15日間培養を行った。」
とある。
細胞のダメージを考慮しての例のようであるが、STAP処理はさらに厳しい酸性に25分さらすのである。こちらもヘテロプラズミーの比率を変化させたであろう。
私もコメント欄でやり取りしているが省く。ここで考え直すこととする。以下昨日の書き込みである。>>
heteroplasmy の違いから - ni -
2020年01月24日
ミトコンドリア病(ヘテロプラスミー)についての情報は近年かなり豊富なようだ。暇に任せ調べていくうちにヘテロプラスミーの比率を変化させるもう一つの知見があった!
それは卵子形成時のボトルネック効果によるヘテロプラスミー割合の変動のみならず胚盤胞(Blastocyst)期でもボトルネック効果があるという。
卵子形成時の早期ボトルネックに対し胚盤胞期のボトルネックは後期ボトルネック(second bottleneack)と呼ばれヘテロプラスミー比率の変動が報告されている。
初期の論文「Rapid Mitochondrial DNA Segregation in Primate Preimplantation Embryos Precedes Somatic and Germline Bottleneck」
イメージ(https://www.lifescience.mext.go.jp/files/pdf/n2132_06.pdf#search=%27blastocyst+bottleneck%27)
例えば、ヘテロプラスミーをもつFES1の株分けらしい129/GFP ESを4倍体胚盤胞にインジェクションし4Nキメラマウスを得る。その4Nキメラマウスを筆頭著者に渡していたらsecond bottleneckによりヘテロプラスミーの比率はばらつきFES1,129/GFP ESとは違ってくるだろうということは容易に想像できる。Aの比率が増えたり(FLS3) Tの比率が増えたり(FLS1) ホモプラスミーになったり(CTS1)...。
エアF1マウスを渡されていたんじゃないか...。
- 2020/01/25(土) 07:46:42|
- ミトコンドリア
-
-
| コメント:0
(あったのか、なかったのか)学ブログはまだ探針を入れているままであるが、最近新しいコメントが入った。
>>
鉛の兵隊
ちなみにFES1がFLS3だとするとFES2は何かということになりますが、おそらくFLS1でしょう。FLSには1月31日に培養開始のものと2月2日に培養開始のもの、2種類あります。FLS3は2月2日培養開始ですが、FLS1は1月31日培養開始です。FLS1はFLS3とは違う仔マウスから作られていると思います。
2020/01/20 URL 編集
鉛の兵隊
>どう作られたかが疑問であるとの問題提起なんです
FES1がどう作られたかは自明です。FLS3と同じだから、FLS3そのものですよ。FLS3を培養してFES1のラベルを貼っただけです。
2020/01/19 URL 編集鉛の兵隊氏は和モガさんだと思っているが、FES2に関してFLS1ではないかという修正説には同意できる。ただし、実証確認もしくは既存実証事実からの演繹はできない。調べられていない。
FES1がFLS3だというのはTs.Markerさんとも一致している。今、Ts.Markerさんのブログに書き込んで回答を待っている。
小保方さんが犯人でない限り、サンプルは若山さんもしくは彼の利害関係者によって中身が入れ替えられているという論理構造になっている。逆に中身の入れ替えがないのなら小保方さんの既存ES捏造だということになる。従って、小保方さんの捏造を証明して警察に突き出さないといけない。
私はそれが可能だと思って、既知の情報から小保方さん犯人説を立証しようとしたができなかった。テラトーマ一つとっても、学生のntESがあるのにGOFマウスがドナーのテラトーマにどうしてFES1をインジェクトするのか。あり得ないことです。しかもESでテラトーマ捏造するのだったら体細胞切り出しなんて不要です。まして、せっかくできのいいテラトーマを捏造したのに3誌段階で一度も使ってない。
大田ESは無かったのにどうして太田ESが使われたということになったのか。太田ESでなくても若山さんの作った129と岡部マウスとのF1ESなんてラボ内にいくらでもあるはずだ。奥さんの論文にも共著者の太田さんのESは使われている。奥さんは保持しているはずです。なぜそんな遠くの京都に取り寄せたのか。ラボには無かったということを示したかったからですよね。ラボにあるFLSもその中の代表ではありませんか。FLSはそもそも129と岡部マウスとのF1マウスから作られているんです。「僕のマウス」を渡したというのが嘘なんですね。近くを調べられると自分の行ったことがばれるから、遠くの太田ESを持ち込んだことにした。そして太田さんの置忘れがあったということを臭わせた。「僕のマウス」を渡したという証拠が何一つない。
細胞の検証は理研の正式な調査チームではなくて、若山さんが率先して行い始めたんです。まず手持ちの細胞を放医研の知人に自分のマウスのコンストラクトと岡部マウスのコンストラクトを教えた上で分析させた。放医研の知人が岡部マウスのコンストラクトの一部が15番の内在性アクロシン遺伝子のプロモーターを捕まえてしまって、GFPの場所を間違えてしまったがために、やはりお友達の遠藤氏の出番となり、間違いを発見したという話を作った。因みに遠藤氏は公共データベースの公開後すぐに分析に着手していて、Kahoの日記で盛んにスピン活動を行っていた。
若山さんはその後、東北大の黒木准教授に太田関連細胞を送った。資金のかかることなので、NHKの藤原記者を巻き込んで、NHKに資金を出させ、東大の知人にも解析させた。その細胞も出所は若山さんである。又、若山さんは理研の松崎と組んで分析を頼んでいる。丹羽さんは途中まで自分で解析していたが、途中から松崎GDにバトンタッチした。そして丹羽さんと相沢さんは小保方さんとともに再現検証実験に回った。その間、理研での解析の中心になったのは松崎GDである。松崎は東大の出身だが理研に来る前は東北大で教鞭をとっていた。
小保方さんの実験ノート3冊分の全コピーがNHKに流出したのは若山さんと松崎グループたちの仕業で、あのNHKの番組に若山さんと遠藤氏、大日向氏が出演しているのは資金援助に対する見返りでもある。須田桃子氏や、古田氏に情報を流しているのも若山さんと理研松崎グループである。査読文は若山さんから出ている。実験ノートのコピーは理研にしかないので松崎GDの違法流出ということになる。公務員法違反であるが、これが不問に付されているのは無論文科省がバックに居て何とかもみ消そうとしているからである。
ただし、若山さんがなぜこんなことをしたのかに関してはちゃんと押さえておかないといけない。若山さんは理研が小保方さんを採用することになる以前の段階で、何一つ公的に悪いことはしていない。ただ、小保方さんとヴァカンティ達に本当のことを言ってなかっただけです。彼にも研究上の秘密を口外しない権利はあります。ヴァカンティ氏との間に駆け引きがあった。
それを世間に出してしまったのが理研の上層部で、しかも笹井さんは信じ込んでいたこともあって、大々的に発表してしまった。誰がどう悪いかという問題はさておいて、理研は若山さんに罪を押し付けることはできないんです。だから何とか玉虫色に鎮静化させたかった。それは文科省との利害とも一致した。そういうことです。
可哀そうなのは小保方さんですが、政治的には他に持っていくことができなかった。それで出版社に手を回して、印税収入が入るようにしてやった。
我々も随分手助けしてあげた結果になっている。
まあ、便所の落書きとしては、事実が分かればそれでいいんで、最後の最後に(あったのか、なかったのか)の問題が残ったということです。
この問題もDORAさんの直感通りあったんですよ。理の導くところあったのでなければ今までの我々の理解はすべて根本からひっくり返って、のみならずどう再構築していいかさえ分からない事態になるでしょう。既に太田ESコンタミは否定されてしまっている。
Ts.Marker さんとの問答で以下のように書いた。
>>
とにかく根本はなぜキメラができたのかという問題です。
①ESによる捏造。
②論文通りにあったのに急遽隠し込んだ。
③ntESによるキメラだということが隠しきれなくなりそうになって①の方向に話をでっち上げた。
②は可能性としてはまだ消えていません。しかし、再現検証実験でどうして簡単にできなかったのか、そしてできたのが真実なら若山さんは記者会見発表までされている段階でどうして突然①だと言い始めたのか、という大きな謎に答えなければならない。
再現には時間がかかると言われている。ムーさんとキンガ・ヴォィニーツさんの論文は再現されましたかね。ムーさんの発明したクレロックスシステムは丹羽さんも再現してますから、これは既に認められているということでしょうね。ただ、筋肉体細胞からの追跡は確認されたのでしょうか。それとキンガさんが行った多能性証明は誰か確認したのでしようかね。これは再現確認されたらノーべル賞クラスではないでしょうか。
分化過程の途中で幹細胞が分化を止めて維持されているという知見は既にありました。筋肉細胞においても筋肉幹細胞がある。ところがこの幹細胞は途中で分化が止まっているのではなくて、一旦体細胞まで分化した後にリプログラムされ、そのリプログラムされた原細胞から筋肉幹細胞が出来ると分かった。しかもその原細胞は体外に取り出してインヴィトロで実験するとキメラが出来るほど深くリプログラムされていると分かった。それが弟子のキンガさんの行った実験ですが、その後をまだ聞いてない。
それに対して、STAP論文で時期を替えて繰り返し何度でもキメラが作られていて、論文の中での再現性は保証されている。しかし、再現検証実験ではキメラができなかった。
これは再現には時間がかかるというような問題ではありませんよね。桂報告書では太田ESによる捏造だったからだという論理になっている。
私はntES化されているからだと答えている。私の言ってることの可能性は桂報告書は考えて居ない。それだけでも既に論理破綻している。考えられる可能性をすべて潰していないから、考え抜けしているわけです。科学者としては超三流でしょ。でもそれは当然で、落としどころとしての結論ありきだからですね。ここもとても良く説明できますよね。超三流の科学者が理研に就職できるのかと考えると分かる。馬鹿なのか人格に問題を持つ悪党なのか、どちらか白状せいと嫌味を言ってる理由です。こういうことからも確信されるわけです。
桂報告書のスタンスは理解可能ですね。我々は第三者ですからね。でも当時者たる小保方さんの怒りは収まらないでしょうね。又ヴァカンティが今後どうするかも未定です。そもそも若山さんも気の毒で、事件化後の行動はともかくとして最初の動機は善意だったんですよね。今、疑われたままになっていて、晴れ晴れとした気持ではありませんよね。玉虫色にするためにたくさんの人たちが犠牲になっている。若山さんも明らかに犠牲者の一人です。
こういうことにしとこうという日本人の悪い面が出てるんですね。何とか丸めたい。そもそも表に出して洗いざらい整理したら世間は普通に納得するということもありますね。誰かが悪いということにしないと収まらないというのは子供心で、社会と言うのは集団活動なので、誰も悪くないのにこうなっちまったんだよというのはいくらでもあり得て、話せばわかるんですけどね。丸めようとする。これは日本人の性癖です。討議するのを嫌がる。昔の意味での大和魂です。盗賊に向かって俺はお前の顔を見たぞとののしったがために盗賊が戻ってきてその人を殺してしまった。なんと大和魂の無い人かと。今昔でしたか小林秀雄が書いてる。
北の拉致者を見て見ぬ振りした。STAP事件も同じですね。処世術なんでしょうね。自分には関係ないと。でも空気に逆らってまでやってもどうせ治らないんだよな。直そうと思ったら一大哲学者の誕生を待たないと無理でしょうけど、今まで出たことは無いね。すると空気に逆らうと損じゃないかということになる。
で、こういうのは積もり積もって国家の危機に結実して行って、そして多分また自分たち自身を犠牲にして特攻し、しかし、敗戦するんだろうな。国破れて山河あり。そして生き残っていればまたガンバルんだ。
じゃあ、どっかにもっとましな国があるのかと言ったところで様々ということなんで、生物は多様性を維持して何かが生き残っていればいいんだということかもしれない。
そろそろ便所の落書きも収束させないといけない。他にやらねばならないことも多い。結局は(あったのか、なかったのか)。ここです。
(あったのか、なかったのか)論理の導くところあったのだ。ただ、実証証拠が無い。
遠藤解析はGOFのFI-SCはGOF-ESと丹羽さんのCD1TSとのコンタミ物だと主張している。
我々は遠藤論文はイカサマだと指摘している。しかし、そこがイカサマであったとしても、GOFのFI-SCが現存してないという桂報告の記載は変わらない。
二つの問題があって両方解けるとあったことを演繹証明できるかな。
①遠藤論文のGOF ESとCD1のコンタミ根拠の検討批判(GOFのFI-SCとCD1のBFPESではないか)
②CTS-1とCTS-11~13の分析結果公開請求。CTS2~10の実験ノート記載公開請求。
私のntES仮説で、このGOFのFI-SCがあったとしたら、これは若山さんの細胞に関する新発見ということになる。
これが分かったからあわてて無かったことにしたというのも考えにくい。こんな事件になってから後から論文書けないね。弟子でも書けないでしょうね。
(ヘテロプラスミーの問題)Ts.Marker さんと討論しているうちに大変な問題に引っかかった。
何よりも先にこの問題を見通さないといけない。(あったのか、なかったのか)は後に回す。
これは和モガさんも、Ts.Markerさんも私も知っている事実だったが、その意味に気づいていないことがあった。今回初めて気づいた。以下のヘテロプラスミーである。
>>
DRR028632 FLS3 T:1949(49%) A:1992(50%)
DRR028633 FLS3 T:1912(49%) A:2015(51%)
DRR028646 FES1 T:3625(80%) A:921(20%)
DRR028638 129/GFP ES T:2963(80%) A:733(20%)
SRR1171585 STAP-SC(FLS?Acr) T:1245(85%) A:217(15%) ??
FLS3とFES1のヘテロプラスミーの割合が違う。これはFES1のチューブにFLS3を若山さんが入れ替えたのだという行為を否定しているように見える。我々は間違ったかもしれない。我々というのは無論私、和モガさん、以前のTs.Maker さん、Ooboeさんとパートナー氏であろうか。
それどころか桂報告書も間違っているということを意味する。桂報告書は遺伝子解析の結果FLS3はFES1だと主張し、小保方さんがFES1を若山さんに渡したから、捏造キメラができたのだと主張していることになる。
そこで我々はその主張はA=Bだということを証明しただけで、小保方さんがFES1を若山さんに渡したことを証明するものではなく、若山さんがFES1の中身をFLS3に入れ替えたのだ、GLSも同様に、学生のntESを小保方さんが若山さんに渡したのではなく、若山さんが学生のntESとして残していたチューブを洗い出してGLSを入れたのだと反論した。そしてどちらが嘘をついているか関して細胞の大きさの違いに気づかなかったといった若山さんの嘘、幹細胞がSTAP細胞より大きくなっていること、GOFのドナーでテトーマに学生のGOF-ESを持っている小保方さんがどうしてF1のESを移植するのだという別の根拠から若山さん犯人説を唱えた。
ところが今新たに気づかれたことは以下の二つの細胞の細胞質内のミトコンドリアDNAのヘテロプラスミーの割合が違うということである。入れ替え云々以前にA=Bではないということらしい。
>>
DRR028632 FLS3 T:1949(49%) A:1992(50%)
DRR028646 FES1 T:3625(80%) A:921(20%)
桂報告書の大本の結論からして違っていたということになる。他がすべて一致しても一つ違っていたらそれは同じものではない。さて、さて。
(ヘテロプラスミーとは)ど素人の知識としてまずブルーバックスの『ミトコンドリアミステリー』を前提しよう。林純一さんの2002年の解説書で2016年15刷版。
ミトコンドリアの中にも二本鎖の環状DNAがある。ミトコンドリアDNA(mtDNA)だ。約1万6千塩基対(bp)で、37種の遺伝子がコードされている。
ミトコンドリアは一個の細胞の中に数千個ある。受精卵の中のわずかに存在している精子のミトコンドリアは卵の中で殺されるので、ミトコンドリアDNAはすべて母性由来である。卵の細胞質内のミトコンドリアだけが次世代へ引き継がれていく。
体細胞分裂時は核のDNAの分裂が終了した後に細胞質が半分に分かれる。その後半分に減ったミトコンドリアの数が倍増されて基の数に戻る。
ヘテロプラスミーというのは数千個あるミトコンドリア内の一つのmtDNAに突然変異が入ることである。数千個の中の1個だけが違うmtDNAになる。異形成と呼ばれる現象。
この現象は体細胞分裂の度に組織内に広がっていく。細胞質は細胞分裂時にランダムに1/2分割されるので一個の変異mtDNAはどちらかに入る。正常な集団はそのままどこまで分割して行っても正常だが、一個入った側は、核分裂の後に倍増されるので2個になる。こちらは次に分裂した時にランダムに1/2されるから一個ずつはいるケースとゼロ個と2個のケースに分かれる。一個ずつ入ると一個入った時のケースに戻る。ゼロ個2個に分かれた時はゼロ個は正常な増殖細胞に戻るが、2個入った側は細胞質内でミトコンドリア数が倍増されるときに4個になる。
こうやっていろんな組織内にヘテロプラスミーが少しずつ蓄積していく。そしてこの広がりは細胞分裂の回数に依るのでいつ変異が入ったかにもよって広がりの程度が違ってくるし、又この変異ミトコンドリアはランダム分割の性質によって変異ミトコンドリアは変異ミトコンドリアばかりに蓄積していく傾向が生じる。
これらは体細胞分裂なので所詮莫大な数のミトコンドリアの中のごく一部なので個体の生存にはほとんど影響しない。問題は次世代にこの異変が引き継がれるとのをどう防ぐかであるが。
卵細胞が作られるとき一旦ミトコンドリアの数が急激に減らされる。例えば千個のミトコンドリアの中に10個の変異ミトコンドリアがあったとする。千個のミトコンドリアを10個に減らす。1/100である。この中に1個の変異ミトコンドリアが入っている確率は1/10である。10個の卵があるとして、9個は正常、1個は変異が入っている。ここから数を元の1000個に戻す。1個の変異の入っている卵は100個入った集団になる。9個が正常に戻った反面、残りの1個には変異がコンデンスされることになる。
そしてこの変異が呼吸に関して深刻な変異であった場合、別の仕組みでこの変異卵はアポトーシスする。つまり自殺するようになっている。これが生殖細胞ボトルネックと呼ばれる仕組みである。これで子孫には正常なミトコンドリアDNAが伝えられる仕組みである。
ただし、致命的でない変異細胞はアポトーシスされないで次世代に引き継がれる。この致命的でない変異も世代によって数が増えてくると致命的になるかもしれない。その時は個体の死によっても排除されていくわけです。この致命的でない変異の問題が今回Ts.Marker さんの発見した変異らしい。
一応以上を前提にTs.Marker さんの発見したChr M 12,188の変異に関して考える。5'末端側の上流から数えて12188番目の塩基が野生型TであるべきところがAになっているという変異である。
しかもわずかに含まれているとかいうレヴェルでなく、何割というレヴェルです。もう一度貼り付けましょう。
DRR028632 FLS3 T:1949(49%) A:1992(50%)
DRR028633 FLS3 T:1912(49%) A:2015(51%)
DRR028646 FES1 T:3625(80%) A:921(20%)
DRR028638 129/GFP ES T:2963(80%) A:733(20%)
SRR1171585 STAP-SC(FLS?Acr) T:1245(85%) A:217(15%) ??
これらはどういう由来であれ培養細胞です。最初に何もヘテロプラスミーの無い状態からどこかで変異細胞ができて、増殖過程でそれが50%になるということは考えられない。培養と言うのは体内で言えば体細胞分裂を繰り返している状態です。一個の細胞には数千個のミトコンドリアがある。培養細胞は一つのシャーレの中に数十万個入っている。この中の一つの細胞の中の一個のミトコンドリアに生じた変異がシャーレ全体の半分を占めるまで増殖するなんてことはありません。
ということはこの変異は生殖細胞ボトルネックをすり抜けた軽微な変異だということになる。
この変異は親の卵にあったのだ。
一応、これらの細胞は裁判レヴェルでの由来検証をされていない、関係当人たちが話しているだけの情報ですが、母親が129/Svだとされている。いまミトコンドリアだけを調べているので、B6は関係ない。
証言が正しいとしましょう。
FLS3は「僕のマウス」の129/Svです。これは「僕のマウス」のロックフェラー大で作ったGFP挿入B6のGFPだけを市販の129/Svに移し替えたものです。雑種にしてからGFP蛍光している子だけに129を戻し交配する。これを繰り返すわけです。3年以上かかる。
この「僕のマウス」の129/Svに以下の割合で一か所だけの軽微なヘテロプラスミーがあったということになるわけです。
>>
DRR028632 FLS3 T:1949(49%) A:1992(50%)
DRR028633 FLS3 T:1912(49%) A:2015(51%)
①これはまず最初から1個の卵の中に数千あるミトコンドリアの半分にヘテロプラスミーがあったと考えることが出来る。
②次にもともとはもう少し少なかったが体細胞分裂過程で増えたと考えることもできる。
③更にTs.Marker さんが見つけた論文にあるように酸浴すると増えるという要素も合わさっているかもしれない。
次に、私のntES仮説であると、移植による129側のヘテロプラスミーがあった場合でも僅かで、大半はICRマウスのミトコンドリアなので、このヘテロプラスミーはこの時に使われたICRのメスにあったものだということになる。以下は上記と条件は同じになる。
①これはまず最初から1個の卵の中に数千あるミトコンドリアの半分にヘテロプラスミーがあったと考えることが出来る。
②次にもともとはもう少し少なかったが体細胞分裂過程で増えたと考えることもできる。
③更にTs.Marker さんが見つけた論文にあるように酸浴すると増えるという要素も合わさっているかもしれない。
次にFES1と129/GFP ESである。
>>
DRR028646 FES1 T:3625(80%) A:921(20%)
DRR028638 129/GFP ES T:2963(80%) A:733(20%)
話ではFES1は太田さんが2005年に作った受精卵ESだということになっているのでまずそれか本当だとする。酸浴はされていない。
①これはまず最初から1個の卵の中に数千あるミトコンドリアの2割にヘテロプラスミーがあったと考えることが出来る。
②次にもともとはもう少し少なかったが体細胞分裂過程で増えたと考えることもできる。ただし、話では目的も無く作られて凍結されているからほとんど継代は重ねられていない。作った時のままであろう。
この話はとてもおかしくて、太田さんはterで作った記憶だと証言している。根本から疑義のある話であるが桂報告書はX1だったという前提で推論している。それが事実であったと仮定して、当時太田さんが使った129/Svには2割のChr M 12,188の変異を有していたということになる。
ここで重要なのは変異は同じ場所だということです。2005年の太田ESに使われた129/Svにこの変異があったわけですが、このマウスが継代して行って5割になるということはありません。どうしてかというと、変異は体細胞分裂によって二分の一ランダム分割によって変異ミトコンドリアばかりが集まっていく細胞が増えていく。そして次世代の卵が作られるときにボトルネック効果で消滅させられる。それでも残った卵で致命的な変異で無ければアポトーシスせずにより濃くなって残るんですが、数的にはずっと少なくなっている。正常な卵の方が多い。これを交配させていますから正常なものが残っていく確率の方がずっと高い。2005年から6年以上ですから3か月ごとに継代したとして24回交配を重ねている。ほとんどいない筈です。
このことによって推測できるのはこのFES1の129は2005年のものではないということです、
DRR028646 FES1 T:3625(80%) A:921(20%)
DRR028638 129/GFP ES T:2963(80%) A:733(20%)
二つは同じ細胞株だとは言えますね。でもこの話は後回しにしましょう。
DRR028632 FLS3 T:1949(49%) A:1992(50%)
DRR028646 FES1 T:3625(80%) A:921(20%)
こちらのふたつの方が大事な問題です。これは同じころの129から作られている細胞です。6年もマウスの形で継代されていたら既に存在してないマウスのはずです。
この二つの細胞が作られた元のメスは共通した変異を持っている。割合が違うだけだ。同じマウスケージの中のマウスのようですね。
上で若山さんの話通りというケースで考えましたが、無論、実際にはこのFLSはアクロシン入りですから、話の通りだとすると、「僕のマウス」の129だとは限らない。若山さんの渡したマウス赤ちゃんは小保方さんによって捨てられていることになっている。そしてFLSの中身はFES1だと結論した。でもヘテロプラスミーの割合が違っていて、かつ、2005年のマウスが変異ミトコンドリアの割合を増やすということは考えられない。となると、FES1はFLSの作成時に近い時期に作られているということになる。
その時の129は何があるのかということです。
>>
①市販129/Sv X1
②①を自家繁殖させているもの
③①を使ってロックフェラー大で作ったB6-CAG-GFPを移し替えたものを自家繁殖させているもの
④市販129/Sv ter (太田氏が好んで使っていたとされるもの)
⑤④を自家繁殖させているもの(これはあったのかどうかは分からない)
④⑤はterです。①は市販時にはTです。②か③しかない。
(培養によるヘテロプラスミーの比率変化)10個の細胞があって、その中の1個の細胞の中の1個のミトコンドリアに変異があるとき、細胞数は10の倍々ゲームで増えていく。10,20.40.80,160…という具合である。そして変異ミトコンドリアを含む細胞は最初は1個で1/10であったが、第一回目の分裂では1/20になる。第二回目では1/40もしくは2/40になる。この原理が分かっていると、シャーレの面積が無限大で細胞分裂がテロメアの長さの許すまで、つまり数10回起こっても、比率は最大1/20で、ランダム分配の在り方によってはもっと小さくなっていくということが分かる。培養では増えない。
ところがシャーレは実際には有限面積なので3日に一回程度セミコンフルになった時点で植え替える。この時、例えば植え継がれた細胞塊の中のヘテロプラスミーが全体の比率と同じであれば、ヘテロプラスミーを含む細胞の全体に占める比率は減りこそすれ増えない。増えるケースと言うのはヘテロプラスミーを含む細胞の集まっているところを偶然に選んだ場合である。この時は生殖細胞ボトルネック効果を人工的に起こしてしまっていることになる。例えば2/40の状態でその2個を含む10個を選んでしまうと、その植え継ぎ時点で、2/10がスタートになってしまう。この偶然が何回か続けばヘテロプラスミーを含む細胞数の割合は増えるし、逆に一度程度なら、継代を続けていればまたむしろ減ってくる原理である。
しかし、今の例は一個の細胞の中に1個の変異ミトコンドリアがある場合の話である。Ts.Marker氏が見つけたヘテロプラスミーはそういうものではない。一個の細胞の中には数千のミトコンドリアがある。
DRR028632 FLS3 T:1949(49%) A:1992(50%)
上の数字の意味するところは全部の細胞を壊してミトコンドリアだけを集めてその中に一か所の同じ変異の有るミトコンドリアが半数だったということなので、これを又細胞膜の中にとじ込めると、変異なしの細胞が半分で、変異ミトコンドリアしかない細胞とが半々にあったか、それともすべての細胞の中が、変異ミトコンドリアと正常ミトコンドリアが半々に入っている細胞であったかのどちらかか、その中間ということになる。
(DRR028652 FES2 T:5307(100%) A:4)ミトコンドリアのヘテロプラスミーと言うのは何種類かの変異が蓄積したもので、この例のようにたった1個の変異だけのヘテロプラスミーがこれほどの割合まで増えると言うのは珍しいのではないか。
特にマウス継代を経て蓄積が増えるのはボトルネック効果を考えてもおかしな話だ。かといって、培養過程で増えるのも原理からして考えにくい。
唯一考えうるのが、Tのホモプラスミーの細胞集団とAのホモプラスミーの細胞集団が混ぜられている場合である。
FLS3は129/B6であるからミトコンドリアDNAは129のものである。129のミトコンドリアDNAの12,188番はTである。
(DRR028652 FES2 T:5307(100%) A:4)はメスの細胞質が違う。
DRR028632 FLS3 T:1949(49%) A:1992(50%)
DRR028646 FES1 T:3625(80%) A:921(20%)
(見かけた論文)>>
その結果, mtDNAのコピー数は最低でも 760コピー程であり, やはり極端なmtDNAの減少は観 察されなかった(Cao et al. 2009). mtDNAコピー数の極端な減少が, ボトルネック効果 の原因ではなかったことから, 他のメカニズムによって ボトルネック効果が起こり, 急調分離を成立させている 可能性が高い.専門家がまだ解に至っていないことに関してど素人は関与できない。既知の確実な知識の範囲内でSTAP事件は解明されるべきであって、それは裁判と同じである。解に至れない場合は分からないとしなければならない。
(Ooboe さん情報)新しい情報が公開されている。ちょっと息抜きしよう。
>>
Ooboe
なにやら、今日もいい文章を構築されてますが、構っていると、焦点分散で前に進めません。真相究明考察、続けます
毎日須田記者は、4月26日作成リスト、5月14日作成リストを早々とリークしてもらった事を、須田著書でけなげに披露しています。この須田記者へのリーク行為から、同様に推察可能なある事が見えてきます。どういうことかと申しますと、、、
2020/01/22 URL 編集
Ooboe
小保方冷凍庫残存サンプル画像リストを毎日須田記者や日経サイエンスなど、早々とリークしたぐらいですから当然、山梨大学若山研にも4月か5月送付していたことは、想像に難くないでしょう。
しかし、石川氏や、新潮記事に登場した理研関係者によると、若山教授やメンバーが紛失した留学生作成ntES78本BOXがリスト画像にあったのに気が付きびっくりした、と証言したのは、理研から公的に送られきた、2014年7月下旬か8月上旬でした。小保方氏立合いで、最終的サンプル帰属が決定しリストが完成した7月19日以後の事です。若山研はすでに、5月には留学生BOX画像リストをCDB解析担当研から入手していたのですから、この7月下旬、8月上旬での、このびっくりした状況を石川氏に証言したメンバーは、石川氏に窃盗印象誘導したことになります。
ところがです、時系列が変なのです。この、7月下旬理研から送られて来た、リストをみて愕然とした若山先生やメンバー達よりも、半月前に、留学生BOXが小保方冷凍庫から見つかって憤慨していた怪人がいたのです。そうです、かの有名な「小保方、地獄の底は、まだまだ、深いぜよwww.」の怪文のCDB内部投稿者。
竹市センター長宛の【引越しのどさくさに箱ごと盗んでいた事を公表しろよ】
2020/01/23 URL 編集
Ooboe
この、若山記者会見の2日後、CDB内部者の怪文内容から読み取れるのは、4月5月小保方サンプル撮影リスト作業した担当者(安全管理室、解析担当研)から、直か、間か、この怪人に内部者でなければ知り得ない情報が流れた事実です。この内容の情報が流れたルーツをたどりましょう。
留学生作成ntES78本BOXを撮影したのは(安全管理室の職員)で手伝ったのが解析担当研ですが、この作業を担当した方々は、若山研が2013年3月ごろCDBから山梨大学への引越し作業には、関わっていません。
ですから、沢山のサンプルの中にあった、このntES78本BOXが、若山研のところから、盗んで来たものなのか?など、分かりようがありません。撮影リスト作成作業をしただけです。
この「引越しのどさくさに若山のところから盗んだ」の怪情報の発信源は、引越しを実際にした、山梨若山研関係者ということに
自然と帰結します。4月か5月にはすでに、CDBリスト作業の解析担研から送付されて来たリストBOX画像を見て「引越しのどさくさに盗まれたもの。」とCDBリスト作業した解析担当研に流していたのが実際のところでしょう。解析担当研、または、関係者に近かったのでしょう。
CDB内部者の怪人投稿は、(小保方盗んだ情報)の発信源の特定に導いてくれました。
7月下旬にBOXがあるのを始めて気付いて愕然としたが如くの、石川氏への教唆証言の虚偽故意性は明白です。
2020/01/23 URL 編集
Ooboe
この怪文投稿のCDB内部者は「丹羽のTsもあったろう」の情報も得ていますから、リスト画像を見たか、
伝聞されたのでしょう。この留学生作ntES78本BOXの「若山のところから盗んだ情報」事案は★2014年5月頃若山研から発信され、
★CDB解析担当研へ、
★そして怪文の投稿者に流れ、
★さらに7月27日NHK偏向番組《NHKスペ》に繋り、
★2015年1月の石川告発に続きます。
この一連の連続性において、CDB解析担当研のメディアを利用したリーク作戦に連携した若山研の動向が鍵になります。この連続性をボカシての、メディア取材の一般論解釈の名文を組み立ててのplus氏解釈は結局のところ、リーク者との特殊実態経緯全体像をはぐらかす、屁理屈反証にしかすぎません。
読みにくいと思います。横書きができませんでした。
2020/01/23 URL 編集
Ooboe
あいかわらず、こじつけが止まりませんがplus氏の情報確認に誤認がありますので正しておきます。
5月14日に作成された当初のリスト表には備考欄はありません。plusさんが日付けを5月14日と思ってしまった、備考欄付きのリストは小保方さん立合い確認の7月19日帰属決定のリスト表のことです。
2020/01/23 URL 編集
Ooboe
リスト備考欄の小保方さんの但し書き「若山研引越しの時に残っていたので保存していた」は2014年7月14日小保方立合いでの帰属決定時点です。
怪文書は6月18日ですので、「引越しどさくさに盗んだ情報」の発信源は山梨若山研に辿りつくのが巻き戻し時系順序です。{BOXが行方不明になって騒いでいたのを知っていた誰かが、リストをみて、怪情報を思い付くことも可能} など、など例によって、しんどい、しんどい、可能性薄いストーリーを無理矢理設定するplus流こじつけは不要です。
引越し作業したのは若山研ですから、5月、送られて来たリストを見て、すぐ反応したでしょう。
それも前後経緯が示す通り、悪意反応の「引越しのどさくさの時、行方不明になってたBOXではないか!」「盗まれていたんだ」と解析担当研に伝える流れになるのが容易に推察できるものです。
回り回って、2015年部外者の石川氏にも、4月26日、5月14日リスト画像をリークしたのも解析担当研、部外者の石川氏がリストを入手しようと思っても、手続きに1ヶ月、しかもまだ公開前。
石川氏は神戸CDBとの直パイプがありません、しかし、同じ横浜理研の遠藤氏に遠藤氏が親しいCDB有力関係者を紹介してもらったのです。そしてリスト等資料と「有志が鍵を付け替えた」証言を教唆され告発を決意したのです。
小保方否定の「若山のところから引越しのどさくさにBOXごと盗んだことを公表しろよ」の、情報発信源とCDB解析担当研との悪意の連携連続性は石川氏へのBOX窃盗告発教唆幇助にまで続いたことで証明されました。
このように、特異な事案が連携連続性をもって存在していた事は、あの姐さんのご指摘は、正論ですがこのケースでは当てはまりません。
2020/01/24 URL 編集
Ooboe
すいません、話の流れで遠藤氏の名義を入れたのは、誤解を招く恐れがあります。不適切でした。お詫びいたします。
遠藤氏は悪意はございません。普通に親切に対応されたとのことです。
2020/01/24 URL 編集
Ooboe
もうひとつ、お断り。
2014年7月19日小保方さん立合い確認でサンプルの帰属決定がなされました。
その時のリスト表は解析担当関係以外は非公開でした。そして石川告発により警察捜査の要請から非公開のままの措置が取られました。ですから、毎日須田記者などメディアでも入手できてません。そうとも知らずに開示請求したパートナーは情報公開担当に捜査終了したら直ちに開示して下さいと手続きしてました2016年6月やっと入手できたのです。
入手できた、情報をある方に伝え、ある方が、Doraさんに、Doraさんが木星さんに進めました
どなたにおかれても、いろんな情報の確認落ちはしかたないですが、頓珍漢の所見にならないようplus氏も気を付けたらいいね、、、
そう言う私も、、、きをつけィ、、です。
2020/01/24 URL 編集理研がー80度フリーザー内に残されていた凍結細胞の内容リストを作成したのが2014/4/5ですね。
撮影とリスト作成作業した担当者は安全管理室と解析担当研なんですね。
小保方さんは入院していましたが2014/7/2に再現実験のために出社した。そしてほどなく自分の-30度冷凍庫内も保全してくれと頼んだ。そして2014/7/19にその両方のフリーザーの内容の口頭説明を行った。
その口頭説明を書き留めたリストをパートナー氏が入手されたのが2016/6月ということですね。その時に2014/4/5の小保方さん説明の書きこまれていない整理リストも入手されましたかね。
桃子本は2014/12/30出版です。巻頭写真にあるリストは小保方さんの説明の無いものですから、最初の整理リストです。桃子は2014/4/5から2014/12/30より前までの間に、理研内の誰かから違法にリークされたリスト写真を手に入れたということですね。
AC129-14に貼り付けているリストの構成は以下です。
①細胞リスト
保管場所 ■■■■■■ -80℃フリーザー [通しナンバー1~23]
②同ボックス中の保管サンプル(リスト以外) [通しナンバー24~27]
③別ボックスの保管サンプル(2014/04/14リスト化) [通しナンバー28~51]
④保管場所 ■■■■■■ -80℃フリーザー [通しナンバー52~152]
平成26年5月14日 確認者 片山 ■■
⑤保管場所 ■■■■■■ -30℃フリーザー [通しナンバー1~55]
確認者 竹市、松崎、丹羽、片山
平成26年7月19日
今ここで問題にしているAC129は②に入っている。(リスト以外)と書かれている。いつ誰が持ち込んだのか。MTA事後締結の際にやり取りが疑われるところです。
まだ続いているようです。
>>
Ooboe
パートナーが広報に3月13日閉鎖という情報がありますから、確認下さいのお願いをしてた件ですが、2014年3月での小保方研究室閉鎖の神戸事業所の作業部署からの報告により、3月18日に変更はないとのことです。
石川調査委員会の記者会見などを確認したそうです。
川合理事との問答があり川合理事の説明要旨は
>「3月13日から、封鎖ではないですが神戸事業所では
小保方研からの移転がなされて行った」
との説明しているのを確認したそうです。
パートナーへの広報の説明では沢山のサンプルがありましたから、封鎖ではないが、出入りを制限しながら小保方研からの、搬出移転保全作業には日数が掛かったのでは、と作業詳細記録がないので、想像のコメントだったそうです。
作業終了報告が3月18日ということは記録にあるので間違いないとのことです。
2020/01/24 URL 編集
- 2020/01/19(日) 11:39:07|
- AC129
-
-
| コメント:2
あらかたの問題は解けた。
幹細胞化の実験は若山さんの研究ですからね。
僕が小保方さんだったらこう言うでしょうね。
私はES細胞でSTAP細胞を捏造してなんか居ないからES細胞に近い性質を持つと書いたと。
ぽこちゃんぺこちゃんブログからOoboeさんは撤退したようだ。私もお気に入りから消した。時間の無駄だ。そもそもあそこは何も新しい情報の無いところだった。照井は血管の専門家だつたかな。道理で詳しいはずだ。素直な心も少しは残っていてチラと出るんだな。はは。TCRのヒントはもらったからな。損はしてない。でももはやああいうことしかできないところまで堕落してるということで、国の研究者は真の発見ができないと教育者になるよりないんだけど、すると人生の目的が違ってしまう。昔デモシカ教師と言われた人々の新版だな。教育は国の根幹で、これはこれでプロの仕事だからな。ここも崩れて久しい。ああいう奴らは愛国心も無いからシナの工作も受けやすくなるということで、今政治でもIR関係で浸透されているのが表立ってきた。米国の圧力だというのが情けないところだが、これからどうなっていくかDORAブログの方が面白いかもね。
同じ、スピン屋でもまだアルイミオウジトゥイッターはフォローしている。あそこにはまだヒントがある。どういう具合にスピンを掛けようとしているかを考えると隠したいことが見える。ぽこちゃんぺこちゃんブログには口汚さしかない。心の中に何か憎しみがあるんだな。妙な奴らだ。国籍が違うのかな。朝鮮総連には成りすましている奴らが多くて、こいつらが実際には日本人誘拐犯だからね。こういうブログではどうせ分からないと思って心の中がさらけ出される。日本人憎しなんだろうな。
Ooboe さんは書き込むところが無くて困まってるだろうね。でもあそこには一般の人はいないよ。一般の人にこの不正を知らせたいなら、まず根本さんブログ、西岡さんガンバレフェイスブックだけどここはスマートフォンからは書き込みにくそうだ。するとやっぱり、パートナー氏のブログ開設が一番ということになる。ガンバレにアップされている動画はもう15万人に増えてる。Ooboe さんたちはこういう人たちに訴えかけたいんでしょ。和モガさんは今ブログを停止しているから相談して、復活させてもらったらどうかな。この間の資料発表会で楠本さんとともに面識を得たんでしたよね。
和モガさんは最初からこの問題を追ってる人だからね。ご自分の説をもう一度纏めて新知見も盛り込んでリヴァイズしてもらうといいですよね。そのシリーズのコメント欄でOoboe さんがサンプルの入れ替えの件を論じさせてもらうといいかもしれない。
Ts.Markerさんは聞いてみないと迷惑かもしれない。ただ、私が延々書きこんでもとりわけ消したりはしないようだ。人は人という感じかな。あの人も言葉不足は有名だからね。でも人は喋れば喋るほど人間が出る。僕は門外だから平気だけど、Ts.Markerさんは一応門内にいる人みたいなんで、あまりしゃべらないように注意しているらしい。このことは以前一研究者ブログで僕に注意してくれたことがあるんで、ああ意識的な行動なんだなと認識している。
それとTs.Markerさんはアルイミオウジ氏を評価している。連絡を取り合ってるようで、流石アルイミさんと言ってる。アルイミ氏はテラトーマにわずかながらOct4-GFPが検出されたのを指摘した人で、最初は無論私は小保方擁護だと思っていた。それとこの発見自体が結構重い事実で、小保方さんがテラトーマを作った時の由来マウスがGOFマウスだという証拠になっている。これはF1マウスは若山さんしか交配許可されていないので他の誰も手に入れられないから当然だと思われているが、それは論理的推論で、アルイミオウジ氏が見つけた事実は物証なんだよな。元のマウスがF1だったということは絶対にないということです。
そのうえでアクロシンを持ち込んだのは誰だ、GFPの無い体細胞は如何にできたのかの問題に解が与えられないといけない。
①無論、私は若山さんがソート前の幹細胞を上から注射したから、リシピエントのES細胞が分化したものだと結論している。
②対して、桂報告やBCA報告は小保方さんが太田ESを使って捏造したと言ってる。そしてGFPの無い組織は免疫不全マウスの体細胞を切り出したと言ってる。間抜けな在米ポスドクに至っては貼り付けて工作したとまでいってる。GLSの捏造には学生にもらったGOF ESが時期的に一致しているのでそれを使ったと言いながら、同じく既に持っていたはずのGOF ESをテラトーマでは使わずに背景の違う太田ESを使ったという。そして小保方さんが捏造したと言っていながら三誌論文までこれを使わなかった理由も答えることができないでいる。小保方さんが出来過ぎを疑ったからに決まっている。
もうこれだけで、桂報告書のインチキは証明されているくらいだ。ただ若山さんが小保方細胞をntES化させたという可能性に気づいていない場合は真相は分からないので、この場合は報告書に分からないと書かないといけない。この段階で既にインチキ報告書になっている。
その上、彼らはntES化の可能性には気づいていたはずである。専門家はキメラが出来るのは既知の知識内ではESかIPSかntESしかないことを知っている。ES捏造を考えてストーリーを作るときに矛盾がたくさんある。その時に彼らはIPSかntESの可能性を考える。特にマウスntESは若山さんが世界の先駆者なんですから考えない筈はない。
ぽこちゃんぺこちゃんブログは桂報告書の構造をそのまま反映しているもので、ESによる捏造説と小保方さん無実説の間に人々を集めようとしている。この中にある限り、ESによる捏造であり、しかし、犯人は分からないというごまかしの中に人々を閉じ込めることが出来ると思っている。そこではntES説は触れられてはいけない説なのである。だからシカト方針になっている。彼らはその可能性を検討しようとはしない。分かり切った話ですね。あの動画を見ようとしている人たちは変だなと考えて居る人たちなんで、ぽこちゃんぺこちゃんブログではだれも納得しないということになる。存在自体が無意味なブログです。
(c,d問題)最後の問題ですね。
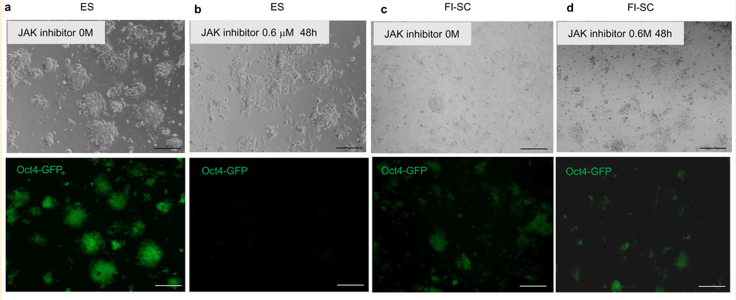
GOF由来FI-SCが存在しなければこの実験はできない。捏造写真は簡単に作れる。さあ、どうですかという問題。ぽこちゃんぺこちゃんブログではシカトされなければならない問題ですね。ぽこちゃんとぺこちゃんの立ち位置の中間のどこにもない問題です。
まずDORAさんの考察から始めましょうかね。
>>
2020年01月14日 12:56
2015年1月1日
じつは遠藤高帆論文もウソだった。
今回の調査報告で、もうひとつはっきりしたことがあります。それは、遠藤高帆論文がでっち上げだったということです。遠藤高帆論文の要点は次の二つでした。
(1)129B6由来の細胞にトリソミーがある。(2)FI幹細胞はESとTSの混ぜ物である。ところが驚いたことに、この二つとも、今回の調査報告では検証されていないのです。まず、トリソミーについてですが、調査報告では、トリソミーはみつかったのですが、それは129B6ではなく、B6の方でした。これでは辻褄が合いません。それから、もっと変なのは、FI幹細胞の方です。遠藤高帆が解析したのはOct4GFPの入ったB6由来のFI幹細胞です。ところが、今回の調査委の報告では、アクロシンの入った129B6由来のFI幹細胞は見つかったものの、
遠藤高帆が解析したB6由来のFI幹細胞は見つからなかった、というのです。?!?!‥‥んなアホな。
遠藤高帆論文を錦の御旗にして「STAPは捏造」と騒いだマスコミ。「STAP細胞・見えてきた実態」とかいう見出しをつけた日経サイエンスの古田彩。このオトシマエどうつけるんじゃ、おんどれ。「遠藤高帆論文はでっち上げでもいいです。忘れてください。今度は調査委員会が別の証拠をでっち上げてくれたので、こんどはこっちを錦の御旗にします」ってか?
これがあんたらのやり口か?結局、遠藤高帆論文の解析結果は、今回の調査報告では全く検証されなかったことになります。遠藤高帆論文は、でっち上げだった。‥‥しかし、問題はそれだけに止まらない。遠藤高帆の解析したFI幹細胞に限って見つからないなんて、ヘンだと思いませんか?他は全部見つかっているのに、これだけ見つからないなんて。これは偶然なんでしょうか? 本当は、見つかったけれども、都合の悪い結果が出たので、隠しているのでないですか? そうとしか考えられないですよ。ますます怪しい調査報告。みなさん、こんなものを鵜呑みにするのですか?
「遠藤高帆が解析したB6由来のFI幹細胞」とDORA氏が言っているのは公共データバンク登録されている以下のデータです。
>>
⑦SRR1171565 若山 FI-SC(Fgf Induced Stem Cells):RNASeq.Rep1 Oct3/4 expressing cells C57BL/6x129/Sv GOF+CD1(丹羽研のBFP-ESの可能性あり)
⑧SRR1171566 若山 FI-SC(Fgf Induced Stem Cells):RNASeq.Rep2 Oct3/4 expressing cells C57BL/6x129/Sv GOF+CD1(丹羽研のBFP-ESの可能性あり)
遠藤論文のTable S1 RNA-seq raw data used in this studyの2P目は以下でした。
上から8,9行目です。確認できますね。
Ts.Marker さんのSRR1171565データは以下でしたね。


このFI-SCは大量にESマーカーを発現し、同時にわずかながらTSマーカーを発現している。2段目のES細胞との違いは明確ですね。これはTru-seq分です。
さて、DORAさんが「じつは遠藤高帆論文もウソだった。」と怒っている遠藤論文の検討です。
>>
Re-analysis of STAP paper: Genotype analysis of fibroblast growth factor-induced stem cells (FI-SCs)
This study examined how SNP allele frequencies in RNA-seq data can be used to show properties of the dataset. Obokata et al. recently reported the phenomenon of STAP, the induced cellular reprogramming of committed somatic cells into pluripotent stem cells that can produce embryonic and placental tissues when injected into blastocysts (Obokata et al. 2014a,b). The allele frequency approach described above was used to examine the NGS dataset provided by the researchers. Allele frequencies between reference allele (equivalent to B6 genotype for dbSNP) and alternative allele (corresponding to 129 genotype in this study) were examined in RNA-seq data from seven replicate experiments obtained using the TruSeq reagent (Figs2A and S1 in Supporting Information). The allele distributions of six of the seven experiments showed the equal representation of parental chromosomes expected in Fig.1A. For the experiments that involved ESCs, STAP cells, and STAP stem cells (STAP-SCs), there were no 0% peaks (Fig. S1 in Supporting Information), possibly because the cells were obtained from mice backcrossed in the laboratory that may have a different genotype than those in the public database (J. Sharif and K. Isono, personal communication).グーグル訳からちょっと手直ししたものです。
>>
STAP論文の再解析: 線維芽細胞増殖因子によって誘導される幹細胞(FI-SC)の遺伝子型解析
この研究はRNA-seqデータの中のSNP対立遺伝子頻度がいかにしてデータセットのプロパティを表示しうるに至るのかを検討している。 小保方らは最近STAP現象を報告した。それは胚盤胞に注入された場合に胚および胎盤組織を作り出すことができる多能性幹細胞へと変化した体細胞の誘導細胞再プログラミングを意味する(Obokata et al. 2014a,b)。上述の対立遺伝子頻度のアプローチは研究者らによって提供されているNGSデータセットを調べてきたものである。参照対立遺伝子(dbSNPのB6遺伝子型に相当)と、代替の対立遺伝子(この研究では129の遺伝子型に対応する)間の対立遺伝子頻度は、TruSeq試薬を使用して得られた7回の反復実験から得られたRNA-seqデータの中で検討されている(サポート情報の図2AおよびS1) 。7回の実験中6回の対立遺伝子分布は図 1Aで期待されている親の染色体と同じ表現を示した。ES細胞、STAP細胞、およびSTAP幹細胞(STAP-SCS)を含む実験では全くの0%のピークは無かったが(サポート情報の図S1)、おそらく細胞が実験室で戻し交配されたため、公開データベース (J. Sharif and K. Isono, personal communication)のものとは異なる遺伝子型のマウスから得られたからであろう。Figure 2-aは以下です。
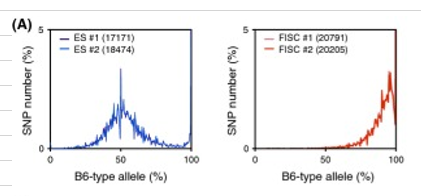 SNPs detected in FI-SC mRNAs indicating contamination. (A) Allele distributions obtained from ESC and FI-SC RNA-seq experiments used in the STAP paper. Both ESCs (blue) and FI-SCs (red) are annotated as having a 129B6F1 genetic background. The number of applied SNPs for each experiment is shown in parentheses in the boxes.
SNPs detected in FI-SC mRNAs indicating contamination. (A) Allele distributions obtained from ESC and FI-SC RNA-seq experiments used in the STAP paper. Both ESCs (blue) and FI-SCs (red) are annotated as having a 129B6F1 genetic background. The number of applied SNPs for each experiment is shown in parentheses in the boxes. Figure S1は以下です。
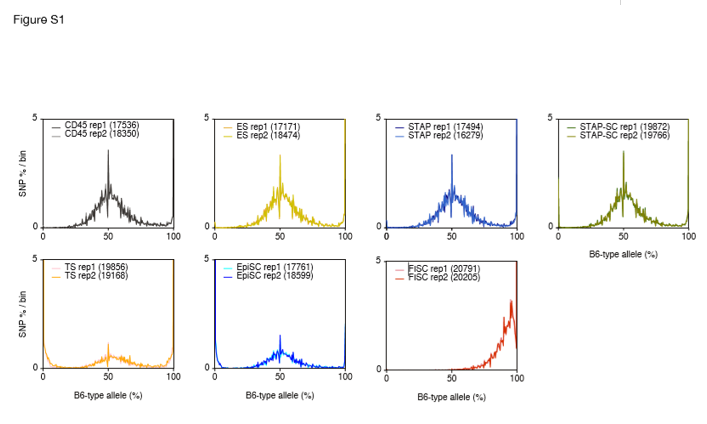
ここで分析されているデータセットは以下です。因みに背景より後ろは後の調査結果の参考メモです。作成者も私のメモです。
>>
Tru-Seq
①SRR1171556 小保方 CD45 positive Cells:RNASeq.Rep1 derived from spleen C57BL/6x129/Sv 129xB6-CAGヘテロ
②SRR1171557 小保方 CD45 positive Cells:RNASeq.Rep2 derived from spleen C57BL/6x129/Sv 129xB6-CAGヘテロ
③SRR1171558 丹羽研 Epi Stem Cells:RNASeq.Rep1 129/Sv
④SRR1171559 丹羽研 Epi Stem Cells:RNASeq.Rep2 129/Sv
⑤SRR1171560 若山 Embryonic Stem Cells:RNASeq.Rep1 C57BL/6x129/Sv
⑥SRR1171561 若山 Embryonic Stem Cells:RNASeq.Rep2 C57BL/6x129/Sv
⑦SRR1171565 若山 FI-SC(Fgf Induced Stem Cells):RNASeq.Rep1 Oct3/4 expressing cells C57BL/6x129/Sv GOF+CD1(丹羽研のBFP-ESの可能性あり)
⑧SRR1171566 若山 FI-SC(Fgf Induced Stem Cells):RNASeq.Rep2 Oct3/4 expressing cells C57BL/6x129/Sv GOF+CD1(丹羽研のBFP-ESの可能性あり)
⑨SRR1171580 小保方 Low pH treated CD45 positive Cells:RNASeq.Rep1 Oct3/4 expressing cells C57BL/6x129/Sv 129xB6-CAGヘテロ
⑩SRR1171581 小保方 Low pH treated CD45 positive Cells:RNASeq.Rep2 Oct3/4 expressing cells C57BL/6x129/Sv 129xB6-CAGヘテロ
⑪SRR1171585 若山 STAP-SC(STAP derived Stem Cells):RNASeq.Rep1 Oct3/4 expressing cells C57BL/6x129/Sv 129xB6-Acr-CAG
⑫SRR1171586 若山 STAP-SC(STAP derived Stem Cells):RNASeq.Rep2 Oct3/4 expressing cells C57BL/6x129/Sv 129xB6-Acr-CAG
⑬SRR1171590 丹羽研 Trophoblast Stem Cells:RNASeq.Rep1 CD1
⑭SRR1171591 丹羽研 Trophoblast Stem Cells:RNASeq.Rep2 CD1
Figure S1を見て直ぐに気づくのは、TSは丹羽さんがCD1のTSを提供したものですからB6タイプアレルなんてあるわけない。若山さんのTSが分化しているようだったから丹羽さんが提供したというのは桂報告が出て後に分かったものですが、データの末尾にCD1と書いてあるのは最初からあるSTRAINの記載です。Epi Stem Cellsも同じで、129/Svと書かれている。で、この原因をbecause the cells were obtained from mice backcrossed in the laboratory that may have a different genotype than those in the public database (J. Sharif and K. Isono, personal communication).と書いていることになる。推測ですね、どんなB6特異的アレルを使ったのかも心もとない。
因みにEpi Stem Cellsは取り敢えず丹羽さん提供と考えているが、使われているのがアーティクル側なので若山さんの作製かもしれない。今AC129に関してここで考えている問題とからんでいる。キメラは作らなかったがEpi Stem Cellsは作ったのかという問題。
ともあれ、Figure 2-aはESは129とB6のF1だが、F1-SCはB6背景だということを示している。ここをDORAさんが「今回の調査委の報告では、アクロシンの入った129B6由来のFI幹細胞は見つかったものの、遠藤高帆が解析したB6由来のFI幹細胞は見つからなかった、というのです。?!?!‥‥んなアホな。」と批判した。
桂報告書が言ってるのは現存しているサンプルとして「B6由来のFI幹細胞は見つからなかった」と言ってるので、このデータベース登録された分は再解析されていて、B6+10%のCD1とされているわけですから、そこは遠藤調査と一致している。というよりもっと細かく調べていることになる。
>>
⑦SRR1171565 若山 FI-SC(Fgf Induced Stem Cells):RNASeq.Rep1 Oct3/4 expressing cells C57BL/6x129/Sv GOF+CD1(丹羽研のBFP-ESの可能性あり)
⑧SRR1171566 若山 FI-SC(Fgf Induced Stem Cells):RNASeq.Rep2 Oct3/4 expressing cells C57BL/6x129/Sv GOF+CD1(丹羽研のBFP-ESの可能性あり)
さて、もう少し遠藤論文の本文の続きを読み進みましょう。
>>
Surprisingly, FI-SCs that were annotated as coming from the F1 129Sv (129) and B6 cell populations did not show the allele distribution pattern of unbiased nonimprinting genes (Fig. S1 in Supporting Information). The distribution was more similar to that of cells with unequal chromosomes. These FI-SCs were reported to be induced from STAP cells with Fgf4 and to have characteristics similar to trophoblast stem cells (TSCs), such as their gene expression profiles and potential to contribute to the placenta (Obokata et al. 2014a).
The obvious difference in the FI-SC curve from the 129B6F1 genotype, combined with the fact that the majority of SNPs were similar to B6, suggested that the FI-SCs originated from neonatal mice of a nearly pure B6 background. Further analysis of gene expression patterns suggested that the heterogeneity of SNPs between B6-type allele and non-B6 could be caused by the expression characteristics of genes. As shown in Fig.2B, SNPs expected to be heterogeneous between 129 (i.e., non-B6) and B6 were examined in several ESC marker genes. ESCs carried alleles from both the 129 and B6 backgrounds at these loci, but the FI-SCs, although described as having the same genetic background as the ESCs (Obokata et al. 2014a), carried only SNPs from B6. This dominance of the B6 genotype was not observed in TSC marker genes (Fig.2C). The FI-SC specificity was not limited to the genes shown in Fig.2B and C. When all heterogeneous SNPs were classified into three groups, SNPs in ESC-specific genes, SNPs in TSC-specific genes, and SNPs in other genes, only FI-SCs had widely heterozygous SNPs for these groups (Fig.2D). If all included cells in a sample share the same cellular features, one would not expect to see this phenomenon of particular gene sets having different genotypes.同様にグーグル訳です。
>>
驚くべきことにF1 129SV(129)とB6の細胞集団由来と注釈されているFI幹細胞はバイアスのないノンインプリンティング遺伝子の対立遺伝子分布パターンを示さなかった(サポート情報の図S1)。分布は不均等な染色体を有する細胞のものにより類似している。これらのFI肝細胞はFGF4<線維芽細胞増殖因子-4 >によったSTAP細胞から誘導され、かつそれらの遺伝子発現の特徴と胎盤に貢献する能力のように、栄養膜細胞(TS細胞)に似た特性を有することが報告されている(Obokata et al. 2014a)。
大多数のSNPがB6と似ているという事実と組み合わせると、129B6F1遺伝子型とFI幹細胞曲線の明らかな差異はFI幹細胞がほぼ純粋なB6バックグラウンドの新生仔マウスに由来することを示唆している。遺伝子発現パターンの更なる分析は、B6型対立遺伝子と非B6間のSNPの不均一性が遺伝子発現特性に起因することが示唆されている。図2Bに示めされているように、129(すなわち、非B6)およびB6間で異質であることが期待されているSNPは、いくつかのES細胞マーカー遺伝子の中で調べられている。ES細胞はこの遺伝子座において129とB6の両方のバックグラウンドからの対立遺伝子を持ち込んでいるが、FI幹細胞は、ES細胞と同じ遺伝子背景を持つと書かれているにもかかわらず(Obokata et al. 2014a)、B6からの対立遺伝子しか持ち込んでいない。 B6遺伝子のこの優勢はTS細胞のマーカー遺伝子には観察されなかった(図2C)。FI幹細胞の特異性は図2B及びCに示す遺伝子に限定されなかった。すべての異質SNPが、ES細胞に特異的遺伝子のSNP、TS細胞に特異的遺伝子のSNP、および他の遺伝子のSNPの3つのグループに分類されているとき、FI幹細胞のみがこれらのグループに広くヘテロ接合のSNPを有していた(図2D)。試料中に含まれるセルのすべが同じ細胞の特徴を共有している場合、異なる遺伝子型を有する特定の遺伝子セットのこの現象は見られ得ないであろう。Figure 2-bです。
 (B) SNPs detected in Sall4 and Klf4, which are highly expressed in ESCs. B6-type alleles are shown in blue and 129-type alleles (i.e., non-B6) are in yellow.
(B) SNPs detected in Sall4 and Klf4, which are highly expressed in ESCs. B6-type alleles are shown in blue and 129-type alleles (i.e., non-B6) are in yellow. 以下のように述べている。
>>
As shown in Fig.2B, SNPs expected to be heterogeneous between 129 (i.e., non-B6) and B6 were examined in several ESC marker genes. ESCs carried alleles from both the 129 and B6 backgrounds at these loci, but the FI-SCs, although described as having the same genetic background as the ESCs (Obokata et al. 2014a), carried only SNPs from B6.
ここは、公共データベース記載の背景にはC57BL/6x129/Svと書かれているが、実際の分析結果は基本B6だと言ってる。小保方さんが登録したんだろと思い込んでいるわけですが、小保方さんは2013/11/5に理研に提出していて、実際にNCBIに登録したのは理研の別の人間で2014/2/13です。
この事件化直後頃に、小保方は公共データベース登録しなければならないことを知らずにいて慌てて2月になって登録したという悪質な噂を流した人間が居て、ここに何か不正がありそうだと誰でも気づいているところですね。
遠藤氏は何かといえば小保方論文に書かれていると書きつけるが、警察でもないのに、何の証拠もなく小保方さんの所為であるかの如くに示唆してはいけない。確かに論文に書かれているが、責任著者には若山さんも居るんだよ。調べてから言わないと。科学者は事実だけを言えばいいんでね。
注に、「(B) SNPs detected in Sall4 and Klf4, which are highly expressed in ESCs. B6-type alleles are shown in blue and 129-type alleles (i.e., non-B6) are in yellow. 」と書かれているんで、ESはF1だがFI-SCはB6だと言ってる。最初に2万箇所前後を統計処理したはずだったが、ここでは全部で10個しか調べてないね。しかもなんだか恣意的な場所だね。小保方さんはESマーカーとしてSall4 なんて選んでないんだから、せめて論文にある9個の中から選べなかったのかな。それだと不都合な真実でもあったのかな。
でも、まあここは理研の後の調査でもFI-SCはB6ベースだったんだからよしとしよう。
10か所選ばれたローカスのどこからどこまでがSall4でどこからどこまでがKlf4なのか分からない。そこで和モガ氏の以下ブログを見ると、一番上のrs33341561はSall4となっているから、それぞれ5か所ずつと推測される。
>>
2017.11.20 Mon l STAP細胞事件 l
「STAP細胞事件」-やはり、「Chip-seq(input) FI幹細胞に欠失はない」でいいんじゃないか
和モガ氏はrs33341561をFLSに導いていって、ほぼ4対6を示すが、遠藤論文はあくまでもESとFI-SCの比較です。それとローカス番号が順に並んでないのはなぜなのか、ど素人には分からない、
和モガ氏の図も拝借しておこう。

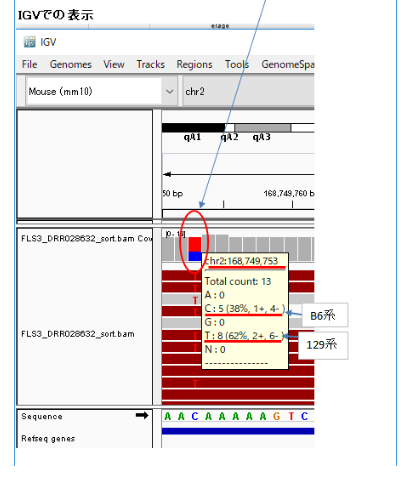
ともあれ遠藤論文Figure 2-bはFI-SCのESマーカーのごく一部のSNPsに関して、これはB6背景マウスだと言ってる。ここでは2万箇所の中の10か所だけが示されているということになる。
次はFigure 2-cだ。
 (C) SNPs detected in the TSC-specific genes Elf5 and Sox21.
(C) SNPs detected in the TSC-specific genes Elf5 and Sox21.bがESマーカーのごく一部を調べたのに対して、対照的に今度はTSマーカーを調べた。
Elf5とSox21を選んでいる。あれほど論文がCdx1を強調しているのに選んでないことはとても不思議です。また、Sox21は後にTSに発現するので研究対象になっていはいるが、TSマーカーとして一般的認識は無かったのではないか。その証拠にESでも出ている。
ただ、ここで意図されているのはFI-SCではTSマーカーとして出ているものの中にわずかなB6以外が出ているということを示している。ただし何度も言うがESマーカーもTSマーカーも10か所ずつしか選ばれていない。全体では2万箇所の統計と自分で書いている。
ESとTSを混ぜたのだという主張の根拠として作られているデータですね。小保方さんが挙げているESマーカーは7種、TSマーカーは3種です。全部示さないといけない。これでは何も言えてない。桂調査はこの混ざったとされる細胞をCD1だと示唆しているが、何を根拠にしているのかを示していないし、混ぜられているという根拠も遠藤調査とは独立して何か示しているかというと何もない。
丹羽さんがESとTSは混ざらないと言ってるのは良く知られている。ここに私が(c,d問題)と名付けた理由がある。あの写真がトリックだったら小保方さんの捏造です。あの実験は理研若山研では行われていません。JAKiの検証をするように指示したのは査読者です。若山さんは既に山梨にいます。
GOFのFI-SCがあったのか、それとも小保方さんがTSとESを混ぜてRNAデータを捏造したのかという問題です。写真はそんなことしなくても捏造なら都合のいい写真を選んで組み合わせるだけですから簡単です。遠藤氏が主張しているのはGOFのFI-SCなんか作られていない。GOFのESと誰かのTSの混ぜ物だと言ってるわけです。
Figure 2-dに移りましょう。
 (D) The number of homozygous/heterozygous SNPs observed in the stem cells used in the original paper. Only the composition observed in FI-SCs would be predicted to affect gene expression. P-values were calculated using Fisher's exact test of genotype distribution between TSC-specific genes and ESC-specific genes. Rep1 and rep2 denote two replicated experiments.
(D) The number of homozygous/heterozygous SNPs observed in the stem cells used in the original paper. Only the composition observed in FI-SCs would be predicted to affect gene expression. P-values were calculated using Fisher's exact test of genotype distribution between TSC-specific genes and ESC-specific genes. Rep1 and rep2 denote two replicated experiments.CD1というのはICRのことで、これは近交系マウスではなくてクローズドコロニーマウスです。これの129特異的SNPだとかB6特異的SNPを調べるという言葉の意味が良く分からない。桂報告書には以下のようにあって、遠藤論文と同じことを書いているように受け止められる。
>>
2)FI 幹細胞の RNA-seq データは二種類の細胞種を含んだサンプルに由来する
(調査結果) FI 幹細胞の RNA-seq データと NGS 解析によるマウスゲノムデータとの比較解析により 得られた SNPs データを詳細に解析すると、大多数のアレルは B6 ゲノム配列と一致する のに対して、5-10%程度のアレル頻度を持つ SNPs 箇所が多く認められる。これは大部分 の RNA-seq データが B6 ホモ系統マウス由来の細胞から得られており、別系統由来を持っ た細胞から取得された RNA-seq サンプルが少量混じっている可能性を示す(少量混じっ ていると考えられる RNA-seq データが示す SNPs 分布は TS 細胞の RNA-seq データ(CD1 系統)と酷似している)。 Figure 2-dのTSCはCD1ですし、EpiSCは129/Svです。図の意図すること自体が理解しにくい。桂報告はGOFに混じっているのはCD1だとまで示唆している。そもそもどうやってそんなことが分かるのか。説明がない。ICR特異的SNPsと言うのはないからね。
以下、和モガさんブログにも書かれている。理屈でもそうなる。
>>
2018.01.29 Mon l STAP細胞事件 l
「STAP細胞事件」-ミトコンドリアの特異点はクローンマウスの痕跡である(3)
一方、クローズドコロニーは 5 年以上外部から種マウスを導入することなく、一定の集団内でランダム交配により維持されている系統で、このため、各個体の遺伝的性質はばらつきがある。従って、マウス系統としてのSNPsを登録できないことになる。問題は遠藤論文ではICRマウスのSNPs解析が実際に行われているということです。裏返すとTS細胞は若山さんの作ったF1のTSだと思っているのではないかという疑義です。遠藤氏はFI-SCに関して公共データベースにF1だと書いてあるにも関わらず、分析結果はGOFだぞ。だから捏造ではないかと言ってる。ということは普通に考えると公共データベースのTSはCD1だと書かれていることを知ってるはずですね。このTSがCD1だと知った上で、それでも129とB6の特異的SNPsで解析することの意味はどういうことになるのか。
まず、勘違いなのか、或はそれはそれで意味があるのかという問題です。彼は以下のように書いていた。
>>
For the experiments that involved ESCs, STAP cells, and STAP stem cells (STAP-SCs), there were no 0% peaks (Fig. S1 in Supporting Information), possibly because the cells were obtained from mice backcrossed in the laboratory that may have a different genotype than those in the public database (J. Sharif and K. Isono, personal communication).
Figure S1ではB6の特異的SNPsを調べているので、少なくとも129/SvとCD1ではゼロピークがあり得るのに、実際には無いからには、ここでは何かにバッククロスされたマウス細胞が使われているのではないかと推測している。
彼は理研の調査チームと違って129/SvのSNPsは調べていない。B6だけ調べていて、B6かB6でないか、B6とB6でないもののF1であるかを区別している。この場合、野生型が入ってしまう。
本文に以下のようにある。
>>
The FI-SC specificity was not limited to the genes shown in Fig.2B and C. When all heterogeneous SNPs were classified into three groups, SNPs in ESC-specific genes, SNPs in TSC-specific genes, and SNPs in other genes, only FI-SCs had widely heterozygous SNPs for these groups (Fig.2D). If all included cells in a sample share the same cellular features, one would not expect to see this phenomenon of particular gene sets having different genotypes.
何をしようとしているのかが示されていないのでチンプンカンプンであるが、まず遺伝子を先に3分しているようだ。
①ES特異的遺伝子
②TS特異的遺伝子
③その他の遺伝子
分析対象細胞は6種。
①ES細胞
②STAP細胞
③STAP幹細胞
④FI幹細胞
⑤TS細胞
⑥Epiblast幹細胞
肝心のマウス特異的SNPsが良く分からない。はっきりしているのはB6特異的SNPsは調べている。しかも、このdの表では2種調べたかのような表示になっている。一応以下だ。
①B6特異的SNPs
②129特異的SNPs
しかし、②は論文本文中では非B6となっている。つまり、B6特異的SNPsで無いものという表現である。129 (i.e., non-B6)という表現がある。B6で無ければ129だという認識だが、
B6特異的SNPのあるローカスにある塩基がB6特異的塩基でなかったとき、それは基本野生型の塩基であって他のマウス特異的SNPではない。129 (i.e., non-B6)の認識は本人が誤解しているのか、スピンを掛けているつもりなのか。
更にホモ、ヘテロの割合を調べている。3種。
①B6ホモ
②B6/129ヘテロ
③129ホモ
正しいのは①だけで、桂チームが行ったように②と③は129特異的SNPsを調べないと分からない。上記疑義したように、彼はB6の特異的SNPがあるとされているローカスの上で、違う塩基があったらそれは129だと誤解しているか、スピンをかけている。これだと③は野生型のホモという定義になる。近交系マウスは無論B6や129の仲間だけではなく、いろんな種類がある。にもかかわらず彼はB6で無ければ129という分類をしている。それは公開データベースに書かれている記載にB6/129と書かれているからなのである。ローカスの間違いと二つに限定するという雑さがミックスされている。
因みに、公開データはこの論文に書かれているように、若山さんがチェックしている。若山氏は背景記述にも責任があるということである。謝辞は慶応の吉村氏で、査読文をばらまいた人である。あちこちにいわば共犯者の名前が散りばめられている。税金に群がる仲良しクラブの構造だということである。
ともあれ、更にすべての遺伝子座が調べられているのではなく、恐るべきことに2百万か所を超えようかというマウスSNPsの中から数個の遺伝子だけを選んでいる。以下である。
①ES特異的遺伝子
↓
②TS特異的遺伝子
選ばれた遺伝子は何なのかの記載もない。例えばEpiSC Rep1のES特異的遺伝子数は190個、逆にTS特異的遺伝子数は87個である。どちらでもない真ん中の遺伝子数は17345個で無論圧倒的に多い。ところがこの圧倒的に多い数の半分しか129ホモではないのである。このEpiSCのマウス背景は129/Svである。
①マウス遺伝子数 約25億塩基対
②近交系マウス特異的SNPs数 約2百万塩基対
③ここでB6特異的SNPsとして選ばれているSNPs数 2万塩基対前後
④幹細胞特異的遺伝子の中でB6特異的SNPsとして選ばれているSNPs数 数10塩基対
捏造論文を疑われても仕方がないレヴェル。もしくは未熟者の書いた論文。
qVALはpVALの誤植ではないか。一番右の欄のp Value は10のマイナス74乗だとか53乗なんて、絶対正しいという確度だが、本当かね。
ESC/TSC>8と1/8の間と言うのは二つの比だけでしょ。どういう恣意的な比較なのかね。ほとんど何を言いたいのか分からない。ただはっきりしている主張はFI-SCはB6背景だということだけみたいだな。雑然とした整理の悪い頭の中だと見える。嘘つきの頭の中も大体こういう構造になっている。
それを確認するために先にFigure 4-bを検討してみよう。

リジェンドは以下である。
(B) Heterozygous SNPs detected in Des, Grb2, Setd7, Fbxo21, and Chd4 where B6 and 129 share the same SNPs but TSCs did not.ここではちゃんと129特異的SNPsが調べられている。ただし、B6と129のそれぞれの特異的SNPの一致しているローカスだけである。つまり
"分かっている"のだ。そして、TSには共通のものが無いと言ってる。当たりまえだろ。それは丹羽研のICRマウスだ。
そして、このTSがFI-SCに1割ほど混ぜられたのだと主張したがっていることになる。ところがまず、TSは全21か所のローカスの中で発現してない遺伝子が9か所あって、分かっているのが12か所そして、FI-SCの中でTSの混ぜ物と示唆されている1割程度の中で一致しているのが12か所である。塩基は4種類しかないので、今、12/21になっているものが12/48であれば単なる偶然ということになる。従って、この一致によってそれがTSの混ぜ物であるということを証明したかったらすべての同様の個所を示さないと有意であると証明できない。ただ単に同じ箇所だけを拾い集めただけじゃないのかと思われて当然だ。
嘘つきなんだな。人格が低劣ということだ。まあ、さもなければ、博士なのに馬鹿ということだ。どっちか選べ。コンチクショウ。
もう一度貼り付けましょう。
2015年1月1日
じつは遠藤高帆論文もウソだった。
今回の調査報告で、もうひとつはっきりしたことがあります。それは、遠藤高帆論文がでっち上げだったということです。遠藤高帆論文の要点は次の二つでした。
(1)129B6由来の細胞にトリソミーがある。(2)FI幹細胞はESとTSの混ぜ物である。
ところが驚いたことに、この二つとも、今回の調査報告では検証されていないのです。まず、トリソミーについてですが、調査報告では、トリソミーはみつかったのですが、それは129B6ではなく、B6の方でした。これでは辻褄が合いません。それから、もっと変なのは、FI幹細胞の方です。遠藤高帆が解析したのはOct4GFPの入ったB6由来のFI幹細胞です。ところが、今回の調査委の報告では、アクロシンの入った129B6由来のFI幹細胞は見つかったものの、遠藤高帆が解析したB6由来のFI幹細胞は見つからなかった、というのです。?!?!‥‥んなアホな。
遠藤高帆論文を錦の御旗にして「STAPは捏造」と騒いだマスコミ。「STAP細胞・見えてきた実態」とかいう見出しをつけた日経サイエンスの古田彩。このオトシマエどうつけるんじゃ、おんどれ。「遠藤高帆論文はでっち上げでもいいです。忘れてください。今度は調査委員会が別の証拠をでっち上げてくれたので、こんどはこっちを錦の御旗にします」ってか? これがあんたらのやり口か?
結局、遠藤高帆論文の解析結果は、今回の調査報告では全く検証されなかったことになります。遠藤高帆論文は、でっち上げだった。‥‥しかし、問題はそれだけに止まらない。遠藤高帆の解析したFI幹細胞に限って見つからないなんて、ヘンだと思いませんか?他は全部見つかっているのに、これだけ見つからないなんて。これは偶然なんでしょうか? 本当は、見つかったけれども、都合の悪い結果が出たので、隠しているのでないですか? そうとしか考えられないですよ。ますます怪しい調査報告。みなさん、こんなものを鵜呑みにするのですか?
2015年1月1日
謎が解けた。
記者会見で桂勲氏が「ES細胞を作製するとき(遺伝子に)かなり変異がはいる」と言ってましたが、だったら細胞の培養中にも変異がはいるのではないでしょうか。「培養変異」として知られています。STAP幹細胞やFI幹細胞の培地は特殊です。そんなところにES細胞を混入したら、まともじゃすまないに決まってます。とくにFI幹細胞の培地では「ES細胞は全滅する」と丹羽さんが証言しているのです。全滅しないまでも、少なくとも遺伝子はダメージを受けて変異が生じるに決まってます。だとすれば、元のES細胞と、培地に混入した後のES細胞で、遺伝子が何から何まで同じってのは、むしろ変じゃないですか。皆さん、調査委の報告が絶対正しいと思い込んでいるもんだから、その結果がどんなに変でも真に受けてしまう。「FI幹細胞の培地ではES細胞は全滅する」っていうのに、「FI幹細胞=ES細胞」だなんて、デタラメですよ。
はっきり言いましょう。調査委の報告書は、何から何までデタラメです。完全なでっち上げなんですよ。それじゃあ、どうやってでっち上げたのかというと、そのカラクリは簡単です。若山研の誰かがSTAP幹細胞とES細胞のサンプルをすり替えて渡したんすよ。最初から、どちらもES細胞ですから、どちらも同じ遺伝子の特徴が現れるのは当たり前です。理研内部にも共謀者がいて、小保方さんの研究室のサンプルは、理研の共謀者がすり替えたんですよ。だとすれば、何もかも説明がつきます。
今回の調査で、遠藤高帆氏の解析したB6由来のFI幹細胞だけ見つからなかったのは何故か。それは、遠藤高帆氏が事前に厳密なSNP解析をしてしまったからですよ。遠藤氏の解析結果は非常に複雑なものでした。TS細胞の混入の可能性もあったし、FI幹細胞それ自体にもわずかにTS細胞の遺伝子の発現があった。非常にややこしかった。
STAP幹細胞は、ES細胞とそっくりの性格を持ちますから、ES細胞とすり替えてもばれる危険性はありません。しかし、B6由来のFI幹細胞は、すでに遺伝子を詳しく解析され、(わずかではあるが)TS細胞と同じ遺伝子の発現を持つという結果を出されてしまった。である以上、犯人は、それと同じような遺伝子の特徴を示すサンプルを用意できなかったのです。ESとTSを適当に混入しても、遠藤氏の出した複雑な解析結果と同じにはならない。話が面倒になるので、「B6のFI幹細胞は見つからなかった」ことにした。これが真相です。
2015年1月1日
謎が解けた(2)
わかりやすく補足します。FI幹細胞は、ネイチャー論文では胎盤を形成する細胞だとされていました。そこで、遠藤高帆氏は、B6由来のFI幹細胞の遺伝子データを解析して、「FI幹細胞はES細胞とTS細胞が9対1で混ざったもの」という結論を出しました。とすれば、今回調査された129B6由来のFI幹細胞の方でも同じような結果が出てもよさそうなものですが、そうではなかった。しかし、なぜ、それに疑問が生じないかいうと、こちらの方は事前に遺伝子解析がなされていませんから、どんな結果でも好きなように言えるわけです。「調査委が出した結果がこうなんだから、こうなんだ」と言われてしまえば、それまでです。しかし、B6由来のFI幹細胞の方は事情が違います。B6由来のFI幹細胞は、遠藤氏によってすでに遺伝子が解析されていますから、それと一致するサンプルとすり替えなければなりません。そんなことをするのは面倒だし、リスクも大きいので、「B6由来のFI幹細胞は見つからなかった」ことにしたのです。そういうことです。
2015年1月2日
調査委の報告が遠藤氏論文と食い違う件。
「遠藤氏の解析は正しいのか、捏造か、どっちだい?」という質問を受けました。正直、私も一瞬混乱しました。厳密に言えば、生データを知っているのは遠藤氏の方ですから、究極的には、私が遠藤氏論文の真偽を判定することはできない。しかし、少なくとも言えるのは、調査報告が遠藤氏論文と一致しないのは、おかしいということです。双方が一致しないということは、少なくとも、どちらか一方がウソを言っていることになります。もちろん、双方がウソという可能性もあります。
いずれにしても、遠藤氏は、「FI幹細胞はES細胞とTS細胞の9対1の混ざり物」という結論を出しました。それなら、調査報告でも、それと同じ結論が出てもいいはずです。ところが、調査報告では、そういう結論を示していません。これは変だと気づくべきです。
ですけれども、調査委の側にも言い逃れをする余地があります。‥‥「遠藤氏が解析したデータはB6由来のFI幹細胞であり、当委員会が解析したのは129B6由来のFI幹細胞である。当委員会が解析したFI幹細胞にはTS細胞の混入は見られなかったのである」‥‥と主張できるわけです。
しかし、仮に、遠藤氏の解析したB6由来のFI幹細胞を犯人がでっち上げるとすれば、どうすればいいでしょうか? 若山研にあるB6のES細胞を持ってきて、丹羽氏のTS細胞と「9対1」の割合で混ぜる必要があります。しかし、そうやって、サンプルをでっち上げても、その遺伝子の解析結果が遠藤氏論文と一致するという保証はありません。たぶん、一致しないでしょう。となれば、非常に話がややこしくなります。ですから、そういう事態を避けるために、「B6由来のFI幹細胞は見つからなかった」ことにしたのです。これが真相だと思います。
2015年1月2日
調査報告は完全なでっち上げである。
今回の調査報告は完全なでっち上げである。以下、その根拠をまとめておきます。
(1)FES1は、かつて若山研にいた人が作製したものである。その人は、小保方さんが若山研にやってくる以前に若山研を去っており、そのさいFES1をすべて持ち去っている。それが若山研に残されていたという証拠も全くない。したがって、小保方さんがFES1を盗むことは出来ない。
(2)したがって、小保方さんの冷蔵庫にあった129GFP・ESは、FES1ではない。
(3)にもかかわらず、「129GFP・ES=FES1」であるという調査委の遺伝子解析はおかしい。⇒これは致命的な矛盾です。
(4)129GFP・ESが若山研で作製されたものならば、若山氏がそれを「知らない」というのはおかしい。誰がそれを冷蔵庫に置いたのかを知っていて「知らない」と言っているのであれば、若山氏はその人物をかばっており、したがって、冷蔵庫にそれを置いたのは小保方さんではない。
(5)若山研に所属するES細胞のケースについて、若山氏が本当に「知らない」のであれば、なおさら小保方さんがそれを「知っている」はずはない。いずれにしても、129GFP・ESを冷蔵庫に置いたのは小保方さんではない。
(6)「FI幹細胞の培地でES細胞は全滅する」と丹羽氏が証言しているにもかかわらず、「FI幹細胞はES細胞が混入して増殖したものである」という調査委の報告はおかしい。
(7)少なくとも、FI幹細胞の培地に混入されたES細胞は遺伝子にダメージを受けているはずであり、それが混入以前のES細胞の遺伝子とピタリ一致するというのは、むしろ不自然である。
(8)遠藤高帆氏が「FI幹細胞はES細胞とTS細胞が9対1で混入したものである」と結論しているのに対して、調査委が単に「FI=ES」と断定したのはおかしい。少なくとも、どちらかがウソを言っている(両方ウソもありうる)。
(9)調査委が遠藤高帆の解析したB6由来のFI幹細胞を「見つからなかった」というのは、きわめて怪しい。おそらく、「遠藤氏の解析結果と矛盾しないサンプルをでっち上げることができなかった」というのが、その真相だと思われる。
以上です。
理研には以下のFI-SCがあった。かつ若山さんはGOF由来FI-SCを作っていないと言っている。
①CTS-1 2012/5/25 Acr-CAG-GFP
②CTA-11~13 2012/7/9 Acr-CAG-GFP
対して、小保方さんはGOF由来FI-SCを作るときに当然GOFマウスを渡されたと思っているし、又、現実にGOF由来FI-SCを持っていたから、後の査読者要請に答えてJAKi検証ができたのである。
この矛盾を解決するのに以下の公開データバンク登録されたデータ解析が行われた。
>>
Tru-Seq
⑦SRR1171565 若山 FI-SC(Fgf Induced Stem Cells):RNASeq.Rep1 Oct3/4 expressing cells C57BL/6x129/Sv GOF+CD1(丹羽研のBFP-ESの可能性あり)
⑧SRR1171566 若山 FI-SC(Fgf Induced Stem Cells):RNASeq.Rep2 Oct3/4 expressing cells C57BL/6x129/Sv GOF+CD1(丹羽研のBFP-ESの可能性あり)
遠藤論文と理研チームの解析では、この細胞のマウス背景はGOFとCD1とされた。
遠藤解析ではTS細胞がCD1であるにも関わらずは129ホモが半分近くあって、データ記載のCD1を失念して、F1であるかのように捏造間違いをしているかもしれない。
理研チームの解析結果は公表されていない。
まず丹羽さんの提供したOct4-GFPとOct4-BFP共挿入ES細胞のマウス背景を確認しないといけない。もしそれがCD1だったらあの実験はGOFのFI-SCと10%のCD1の混合で矛盾がなくなる。
もっとも、いずれにせよ、GOFのFI-SCはあるかないかのどちらかでなければならない。
DORA氏の疑義はまだ解決していない。
- 2020/01/14(火) 22:35:36|
- AC129
-
-
| コメント:0
さて、STAP問題が忘れ去られないように周りを掻きまわしていたら、自分の本題を思い出せなくなってしまった。
AC129の実験とは何であったかということでした。我々は小保方細胞核使用ntES論者です。小保方さんを引き留めるためにntES化によって作成したキメラをSTAP細胞本来の力でできたかのように一時的に嘘をついた。ただ、実験そのものは小保方さんを山梨に連れて行けたら研究課題にしようとしていたもので、真面目な研究だった。翌年に助手で誘ったが返事を保留されてしまって、本当のことを言えなくなってしまっているうちにヴァカンティが本気になって米国の特許仮申請をしてしまった。無論、後から何かの間違いだったと言い訳することは出来るので、しばらくは自分の実験に熱中していた。そして胎盤が光るということを見つけた。この間に小保方さんの論文は3誌にリジェクトされる結果になった。その途中で西川さんのアドヴィスを貰って、TCR再構成の検査を行ったが、幹細胞とキメラのPCR解析結果に自分の行ったntES作製ではあり得ない結果が出たという疑義があったのではないかと推測して見ている。その後にAC129の実験を行ってる。キメラができたローザだと小保方さんは聞いているが、後に捏造だとされた。この流れが良く分からないのであった。
(特許のキメラTCR再構成バンド)①桂報告書ではこの実験はマウス背景の違いによってSTAP細胞のでき方にどんな差があるかを調べる実験とされている。
②小保方さんは渡されたマウスを129/Sv carrying Rosa26-gfp だと書いている。この場合若山さんがなぜこんなマウスを渡したかと考えると、細胞のトレースのためだとしか考えられない。
③①にせよ②にせよキメラまでは当然作らないと差は分からない。
④若山さんは①だと言いながらキメラは作って無いと言ってる。作っていたのに廃棄したからだと考えるのが普通で、キメラがあると背景が分かる。幹細胞なら中身を入れ替えられる。
⑤問題は何をどうやってトレースしようとしたのか。
⑥そして、なぜ、トレースしてみようと思ったのか。
⑦ムー論文は既に出ていてトレースの仕方は分かっていてる。丹羽さんがやったのと同じことだ。これでクローン胚を作ってntESにしてみたらどうなるか。
⑧キメラが出来ることは当然なのだから、問題は胎盤が光るかということになる。
⑨この頃若山さんは自分が普段使っているntES由来の胎盤は光らないと思っていて、小保方細胞核使用ntESから作ったキメラ胎盤は光ると思っている。だからこそFI-SC誘導細胞を作ろうとしていた。
⑩このローザの実験でのキメラ胎盤も当然光ったことになる。仮に見間違いだとしても、同じように見間違うのだから胎盤に関しては光ったと確認できたことになる。
⑪白血球の時はT細胞に当たるか否かの保証がなく、何がSTAPになったか証明できなかったが、このローザクレマウスは肝臓の細胞だと分かっている。キメラは出来て当たり前だが胎盤も光っている。今までのntESの胎盤は光らない。小保方細胞核を使ったから光ったのだという結論になる。この場合キメラも幹細胞も129/Sv carrying Rosa26-gfp だということになる。
⑫しかし、ある時自分はずっと勘違いしていたと気づいたとしよう。そしてそのことを誰にも知らせまいとするとき、幹細胞の中身は入れ替えて小保方さんのESによる捏造にしないといけない。129のESは小保方さんは持ってない。だから「僕のマウス」ESを入れた。キメラは129でできているので「僕のマウス」キメラには色が違うからできない。だから捨てて、キメラは作って無いと言った。
⑬ここでの問題は何時どうやって気づいたかということになる。
⑭レター論文の笹井さんの追加実験結果かもしれない。
⑮もしそうなら、引っ越しの時の細胞の入れ替えのみならず、MTA再締結の頃、つまり石川フライデーのカギの付け替え問題とも絡んでいるかもしれない。
⑯ではレター論文のどの実験で若山さんは自分の勘違いに気づいたのか。
⑰若山さんは8月頃責任著者を降りたいと笹井さんに言った。この頃にはJAKi検証結果が出ていて、寺下さんからその情報を得ている。木星リストに2013/5/14及び15と書かれている。
⑱そんな検証方法があるならと、今までの自分のGOFのntESをJAKi検証してみたら蛍光が消えなかったので、そもそも自分の作っていたntESは通常のESとは違う性質を持っていたのだと気付いたのかも知れない。小保方細胞は関係ないのだと。
(記録)前のブログで以下のように書いた。
>>
(Ooboe さんへの連絡)
まあ、冗談はさて置いて、Ooboeさんたちは自分のブログを早く開設した方がいいと思いますよ。そしてTs.Marker さん、和モガさん、根本さん、DORAさん位に知らせて、そこで自説展開して意見を求めていたら、人々が集まってきますよ。私にも知らせてくれたら伺います。その内興味のある人たちが読むようになります。
書き込む場所がないから学さんのところに間借りしているんでしょ。僕はあそこはため息氏と学氏はほぼ同一人物だと思ってますよ。同一チームかな。そこにいくらOoboe さんが書き込んでも悪口の対象になるだけで、世間の一般人は読んでませんよ。あれは興行手段です。珍しくないじゃないですか。
ため息なんてしたらば荒らしの犯人じゃないですか。金曜日にいつも出掛けている。ああ、これはジムさんしか知らない話でしたか。
パートナーさんに是非ブログ開設してもらってください。難しくないよ。何なら若い人に作ってもらったらいい。本日学ブログに以下の書き込みを行ったら、後で3と4を消された。学さんは照井さんのようです。本当に困るんですね。>>
学さんへ1
あなたは開業医をされていたんでしたっけね。医者だということはご自分でおっしゃってたが、何か不注意でため息グループに暴き出されたのでしたかね。そして何かホームページにいたずらされた。当然ですが、それを怒ってらっしゃった。
お気持ちは分かりますが、あちらはああいう仕事が好きな人なんでしょ。パソコンいじりばかり書いてる。僕なんかああいうのは音痴なんで逆にうらやましいくらいですね。財務省出身の異色経済評論家の高橋洋一氏なんか自分でパソコン作るとネット番組で自慢してる。あの人は東大の数学でしたよね。まあ、理系の人は仕事でもあるんで、基礎から学んでますよね。
我々は仕事の関係で必要最小限の取説だけ学んで使う。ああいうのは面白そうで興味はあるんだけど、中途半端にやっても面白くないし、趣味で勉強するには他にやりたいことが多すぎるということで、ただ羨ましがってるだけということになってしまう。あはは。
学さんへ2
でも、悪事にそれを使ってると話は別ですからね。違法行為なら警察に届ければいいですね。違法とまではいかない場合はそれは法的な解決の無い場所ですから、目には目を歯には歯をのハムラビ法典でいいんです。やられたらやり返せばいい。まあ、復讐するときは10倍返しです。やったらやり返されるのだと悪人に知らしめないと悪弊が横行してしまいますから、それこそが社会正義だと言っていいでしょう。体育館の裏に呼びつけといてボッコボコに蹴り入れてやればいいんですよ。お前どこで転んだんだと笑ってやればいい。
ただし、警察に届けられたら取り締まられるような範囲まで出てはいけない。相手はそこまでやってないんでしょ。やってたら警察に届ければいいだけで、ちゃんと法が裁いてくれる。悪事と言うのは法の網の目をすり抜ける範囲でしか行えないんです。そこが道徳と違うところで、正常な人間は普段意識してないが道徳律に縛られていて、法律よりずっと細かい網の目の中で生活している。悪人と言うのはみんなが道徳的なのを利用して自分の利を図るんですから、逆にみんなが道徳的であることを止めると全然自分の利を図れなくなってしまう。現在の中共の騙しあいのシナ社会がそうですね。日本はあんな野蛮国ではない。
学さんへ3
>>
本人が希望しない状態で
私のため息ブログを見るところご本人は所謂"出臍"です。目立ちたがり。クラスに数人必ずいるでしょ。大人になって芸能人になったりする子。Terui先生はそこまでの、つまり、抑えきれないほど内から湧き出る"出臍"の才能は無いんですね。だから芸能人にはなれなくて学者どまりなんです。
それと彼は隠れている気なんてさらさら無いと見えます。というよりあのブログの目的は学内で何かの連絡に使ってるものではないですかね。Teruiと書いてあるから、あのブログを利用している人たちはあれはterui先生だと分かっている。多分学内ですから書いてなくても関係者が分かっているくらいではないですか。勤務先の大学は目白大学でしたよね。自分で書かれているんですよ。学さんがこれはいけないと思われるなら○○大学に直されて構いませんが、それって逆に○○大学に失礼じゃないですかね。彼は学内で他の先生を批判したりしている。でもそれはご本人の自由です。学内で批判もできないで学問の自由なんてあったものじゃない。ご本人は教員で、しかも博士さんですからね。逃げも隠れもできない仕事で、いつもその身を律していないといけない。我々一般民間人とは立場が違います。彼は何にも悪いことしてないから堂々と自分の名を出しているんです。私の見るところでも彼にはやましいことは何も無いと見えます。学先生はお医者とは言え、開業医でしょ。民間人だ。学校の教員ではない。価値観は同じではないですよ。
学さんへ4
学先生はteruiさんのブログで自分の病院のことをいろいろ書かれ、悪口を言われ、ババア(失礼)呼ばわりまでされていて、悔しいんでしょ。でもヤフーに言っても警察に届け出ても解決できませんよね。なぜかというと法の網の目の大きさの範囲内だからです。そういう場合はどうしたらいいかというと、、、ハムラビ法典でしたよね。10倍返しです。体育館の裏に呼びつけて蹴りを入れてやればいいだけです。喧嘩するときは自分の有利な場所で行う。戦争と同じです。鵯越の逆落としです。背後を襲う。体育館の裏に仲間と待ち伏せして置く。戦争はなんでもありです。女だからそういう発想をできないんですよね。まあ、裏返すと女性を相手に彼が如何に女の腐ったような男かということです。
学さんへ5
で、私のため息ブログを子細に読んで鑑みるところ、とても立派なことが書かれていて、terui先生はそういう"女の腐ったような男"には見えないんですけどね。これってとても不思議なんですけど、唯一、理解可能なケースとしては、私のブログに書いているように、terui先生とgaku先生は同一人物だと思えば、別に自分自身にババア呼ばわりしたところで何も"女の腐ったような男"だということにはなりませんよね。
所謂マッチポンプですけど、どんな世界でも常套的に使われている手法で国際間の外交なんかでは当たり前ですね。
でも、私はそんなことはどうでもいいんで、真実を知りたいだけですから、私の方針であちこちに書き込むんです。無論、自分のブログにもです。すべて私の判断で他人の判断の入る余地はありません。
私はしたらば掲示板を荒らされましたが、あれはKCIA出身の半島人の経営している情報収集用ネットなんで、分かって書いてますからどうなっても構わないんですが、荒らした奴は別人ですので、復讐はいずれする予定ですが背後を襲いますからここでは分からないでしょうね。そいつの生活に実害を与えたらいいだけですからネット上でやる必要はない。でも、ウクライナ航空みたいに誤爆するとまずいですよね。慎重に判断しないと。
学さんへ6
>>
あちらの皆さん、どの方も進化がないじゃあないですか。同じスタンスで同じ事しかいってません。
文科省と関与したNHKを含むマスコミの雇われ人と若山さんのお仲間グループによるスピン工作屋達なんですから当たり前ですよね。わずかな小遣い貰って乞食生活しているんですから、進化なんてする気のある筈はないじゃないですか。進化したら学さんがあの花咲かアルツハイマーお爺さんに小遣い上げるんですか。あなたはそんな相手といつまでも言い争っておられるから、ああ、terui さんだと私なんかに思われてしまうんでしょ。
どうですか。学さん。私の妄想を打破してやってください。
彼らを相手にしないでザハリッヒに問題を考えませんか。
Letter Extended Data Figure 5-c,dは本物ですか、それとも小保方さんの画像捏造ですか。私は一度ため息ブログに書き込んだことがあるんですが、彼は学さんと同じように何か人のコメントを切り取ってアップするんですよね。そして最後にはあなたはあなたでやってくださいと言いましたかね。それ以来連絡はしませんが、学さんが小保方擁護を装って人を集め、ため息ブログで叩くという手法ですから、学さんはいつも論破される側の藁人形役です。ときどき私なんかが出て行くと、コメントを遮るんですね。あれは照井本人です。
まあ、成程そこまでやってるのかと理解すると、ES説は嘘なのだと逆に確信が持てますね。彼らの存在自体が証明している。彼らが居なければ自然に忘れられていくと思いますけどねえ。便所の落書きしているのは私くらいのものだということでいいのにねえ。それがそうならないのは彼らは恐れているんですね。内心の疚しさに耐えられないので、いつまでも見えない敵を見張っている。天知神知我知子知。忘れられないんだな。ひどいことしちまったなあと。
我知。(メモ)(中立的STAP関連ブログ)
❶小保方晴子さんへの不正な報道を追及する有志の会 [2018/3以降完全休眠中]
❷和モガブログ [2018/11以降完全休眠中]
❸和モガトゥイッター [2018/11以降完全休眠中]
❹アトモス部屋 [ヤフー終了に伴い消去]
❺Ts.Markerブログ [ヤフー終了に伴い消去]
⑥Zscan4のブログ [TSさん引っ越し]
⑦Ts.Markersトゥイッター [平常運転]
⑧DORAブログ [引っ越し後平常運転:STAP問題は基本降りている]
⑨一研究者・教育者の意見 [2020/1に2年ぶりゴーン氏ネタで復活]
⑩根本ブログ [引っ越し後現在ヴァカンティの特許のみフォロー中]
⑪がんばれ、小保方晴子先生!西岡フェイスブック [平常運転]
⑫小保方晴子さんの病気が完全回復し、毎日を笑顔で過ごせるように [ジムさんのブログ:平常運転]
⓭ryobu-0123のブログ [2018/2以降完全休眠中]
(分からあああん奴)
⑭阿塁未央児トゥイッター [平常運転:TS氏評価]
(工作員系)
⑮「結論ありき」の非専門家ブログが社会を混乱させている:世界三大不正STAP事件の正しい理解を社会に広める会 [2018/11以降完全休眠中:雑談コーナーのみ平常運転:感想のブログ-元若山研のテクニシャン]
⑯ため息ブログ [平常運転:照井氏のブログ]
⑰考察学とみ子 [平常運転:照井氏の成りすましブログ]
⑱学とみ子のブログ [平常運転:照井氏の成りすましブログ]
(DORA氏ブログへの書き込みメモ)2019年12月28日
小保方さんが結婚してた?
コメント
1. 一言居士
2020年01月14日 12:37
彼は忙しいらしくて愛想無いので、ここに置手紙して置こう。ひひひ。
私にとっての問題意識のあるところ、他は彼から学んだし、現在共通認識なので触れない。
2014年4月4日
もしSTAP細胞が捏造だったとすれば(2)
>>
(調査委員長の石井俊輔氏は「なぜこのまま発表しなかったのか」と言っている)
読み手が論文論旨を理解しやすく提示するのが図の役目と認識している。それが捏造を生みやすいとか、科学的に自分の認識の間違いを誘発しやすいから、ルールがあるということを教えられてない。
2. 一言居士
2020年01月14日 12:39
2014年8月28日
CDBは解体した方がいいのでは?
>>
研究者なら誰でも知っていると思うけど、笹井氏レベルの科学者っていうのは、ほんのごく一握りで、あとはほとんどクズ。重箱の隅をつつくようなことばかりやっている。ただ、それを言うと9割の研究者が失職するので誰も口に出して言わないけどね。
誰が将来の笹井氏か事前には分からない。民間企業の研究所では千三つの当然のコストとして処理されている。
3. 一言居士
2020年01月14日 12:42
2014年9月21日
若山研、ES細胞移管手続きの書面が存在しない?
理研上層の指示。事務手続きはマニュアル化されているので忘れるとかうっかりはない。発表までの守秘のため。
2014年9月26日
捏造の証拠を捏造する。
>>
遠藤高帆氏がトリソミーの一件を論文にして発表したらしいが、これにもおかしな点がある。なぜなら、若山氏所持のSTAP細胞サンプルにも、理研が分析したSTAP細胞サンプルにも、トリソミーはなかったからである。
遠藤の解析間違い。
4. 一言居士
2020年01月14日 12:43
2014年12月23日
あらためて「自家蛍光」説(3)
>>
そして若山氏は、「研究室のスタッフが小保方さんの隣でじっくり見て、メモをとって、それを忠実に繰り返しても、なぜか成功しないのです」と証言している。
ESを混ぜたと誘導している。
2014年12月23日
捏造の証拠はいまだに一つもない。
>>
真葛原雪氏によれば、ゲノムインプリンテイングのある哺乳類では、アリル特異的な発現があるという。つまり、一方のアリルだけが偏って発現することもあるわけで、あながちトリソミーが原因であるとは言えない。
>>
「FI幹細胞=ES+TS」というのもおかしい。なぜなら、TS細胞に特有の遺伝子の発現に、B6側からの寄与があるからである。このことは、「FI幹細胞」にもともと胎盤形成能力があることを示している。私は遠藤論文は怪しいと思ったが、やはりというか、「FI幹細胞の培地ではESは増殖しない」という丹羽氏の証言を得た。
遠藤論文の論証の粗雑さ。データの証拠能力調査なし(公共データベース登録マウス背景記入は他者が関与している)。
5. 一言居士
2020年01月14日 12:45
2014年12月30日
やはりSTAP細胞は存在する。
>>
マウスは成熟するのに3ヶ月ほどかかります。「世代を超える」のに十分な期間がありません。逆位反復配列を維持したマウス集団を共有すれば、STAPとESの双方に逆位反復配列が現れる可能性はあります。しかし、これはあくまで可能性として論じているだけで、どちらの可能性が大きいかといえば、やはり混入の可能性の方が大きいと思います。
>>
AC129とFLS-T1,T2は、この中のES1とだけ、きわめて酷似しています。四つの遺伝子の欠失、SNPタイプ、性別、何からなにまで同じです。こういうことは、まず偶然ではあり得ませんから、これは混入だと思います。
若山さんによる中身の入れ替え。「僕のマウス」ES1は山梨の若山さんが最終的に持っていて、理研には存在してない。
6. 一言居士
2020年01月14日 12:50
2014年12月30日
STAP細胞の存在を証明してしまった調査委員会。
>>
調査委は、残されたキメラ子やテラトーマの遺伝子を解析していますが、それらはすべてFES1由来であると結論していることです。しかしながら、実験当時、FES1は若山研に存在していなかったのですから、それがFES1であるはずがありません。ということは、その正体は、それと同じ特徴をもつFLS3だったということになります。STAPは存在したのです。墓穴を掘りましたねえ。
STAPは小保方細胞核使用ntESとして存在し、そのままFLS誘導したとされた。これだと動機の説明が可能。
2015年1月24日
「一研究者・教育者の意見」に一本とられたのだけれども。
>>
「小保方氏自身も捏造と認めたDNAのメチル化のデータについてである。STAP幹細胞がES細胞であったとしたら、データを改ざんする必要はなかったはずなのだ」
その通り。
7. 一言居士
2020年01月14日 12:53
2015年1月9日
「99.95%一致する」について
>>
スライドの表には「FES1とFES2で異なるSNPsのみ使用して比較」とあります。そんなことをして何の意味があるのかわかりません。これでは、FLS3とFES1は「同一マウス由来」ということの証明にしかなりません。
その通り。ここまではちと気になったことだけ。ここから先はDORA説の全文。コメントなし。
8. 一言居士
2020年01月14日 12:56
2015年1月1日
じつは遠藤高帆論文もウソだった。
今回の調査報告で、もうひとつはっきりしたことがあります。それは、遠藤高帆論文がでっち上げだったということです。遠藤高帆論文の要点は次の二つでした。
(1)129B6由来の細胞にトリソミーがある。(2)FI幹細胞はESとTSの混ぜ物である。
ところが驚いたことに、この二つとも、今回の調査報告では検証されていないのです。まず、トリソミーについてですが、調査報告では、トリソミーはみつかったのですが、それは129B6ではなく、B6の方でした。これでは辻褄が合いません。それから、もっと変なのは、FI幹細胞の方です。遠藤高帆が解析したのはOct4GFPの入ったB6由来のFI幹細胞です。ところが、今回の調査委の報告では、アクロシンの入った129B6由来のFI幹細胞は見つかったものの、遠藤高帆が解析したB6由来のFI幹細胞は見つからなかった、というのです。?!?!‥‥んなアホな。
遠藤高帆論文を錦の御旗にして「STAPは捏造」と騒いだマスコミ。「STAP細胞・見えてきた実態」とかいう見出しをつけた日経サイエンスの古田彩。このオトシマエどうつけるんじゃ、おんどれ。「遠藤高帆論文はでっち上げでもいいです。忘れてください。今度は調査委員会が別の証拠をでっち上げてくれたので、こんどはこっちを錦の御旗にします」ってか? これがあんたらのやり口か?
9. 一言居士
2020年01月14日 12:57
結局、遠藤高帆論文の解析結果は、今回の調査報告では全く検証されなかったことになります。遠藤高帆論文は、でっち上げだった。‥‥しかし、問題はそれだけに止まらない。遠藤高帆の解析したFI幹細胞に限って見つからないなんて、ヘンだと思いませんか?他は全部見つかっているのに、これだけ見つからないなんて。これは偶然なんでしょうか? 本当は、見つかったけれども、都合の悪い結果が出たので、隠しているのでないですか? そうとしか考えられないですよ。ますます怪しい調査報告。みなさん、こんなものを鵜呑みにするのですか?
10. 一言居士
2020年01月14日 12:58
2015年1月1日
謎が解けた。
記者会見で桂勲氏が「ES細胞を作製するとき(遺伝子に)かなり変異がはいる」と言ってましたが、だったら細胞の培養中にも変異がはいるのではないでしょうか。「培養変異」として知られています。STAP幹細胞やFI幹細胞の培地は特殊です。そんなところにES細胞を混入したら、まともじゃすまないに決まってます。とくにFI幹細胞の培地では「ES細胞は全滅する」と丹羽さんが証言しているのです。全滅しないまでも、少なくとも遺伝子はダメージを受けて変異が生じるに決まってます。だとすれば、元のES細胞と、培地に混入した後のES細胞で、遺伝子が何から何まで同じってのは、むしろ変じゃないですか。皆さん、調査委の報告が絶対正しいと思い込んでいるもんだから、その結果がどんなに変でも真に受けてしまう。「FI幹細胞の培地ではES細胞は全滅する」っていうのに、「FI幹細胞=ES細胞」だなんて、デタラメですよ。
はっきり言いましょう。調査委の報告書は、何から何までデタラメです。完全なでっち上げなんですよ。それじゃあ、どうやってでっち上げたのかというと、そのカラクリは簡単です。若山研の誰かがSTAP幹細胞とES細胞のサンプルをすり替えて渡したんすよ。最初から、どちらもES細胞ですから、どちらも同じ遺伝子の特徴が現れるのは当たり前です。理研内部にも共謀者がいて、小保方さんの研究室のサンプルは、理研の共謀者がすり替えたんですよ。だとすれば、何もかも説明がつきます。
11. 一言居士
2020年01月14日 13:00
今回の調査で、遠藤高帆氏の解析したB6由来のFI幹細胞だけ見つからなかったのは何故か。それは、遠藤高帆氏が事前に厳密なSNP解析をしてしまったからですよ。遠藤氏の解析結果は非常に複雑なものでした。TS細胞の混入の可能性もあったし、FI幹細胞それ自体にもわずかにTS細胞の遺伝子の発現があった。非常にややこしかった。
STAP幹細胞は、ES細胞とそっくりの性格を持ちますから、ES細胞とすり替えてもばれる危険性はありません。しかし、B6由来のFI幹細胞は、すでに遺伝子を詳しく解析され、(わずかではあるが)TS細胞と同じ遺伝子の発現を持つという結果を出されてしまった。である以上、犯人は、それと同じような遺伝子の特徴を示すサンプルを用意できなかったのです。ESとTSを適当に混入しても、遠藤氏の出した複雑な解析結果と同じにはならない。話が面倒になるので、「B6のFI幹細胞は見つからなかった」ことにした。これが真相です。
12. 一言居士
2020年01月14日 13:01
2015年1月1日
謎が解けた(2)
わかりやすく補足します。FI幹細胞は、ネイチャー論文では胎盤を形成する細胞だとされていました。そこで、遠藤高帆氏は、B6由来のFI幹細胞の遺伝子データを解析して、「FI幹細胞はES細胞とTS細胞が9対1で混ざったもの」という結論を出しました。とすれば、今回調査された129B6由来のFI幹細胞の方でも同じような結果が出てもよさそうなものですが、そうではなかった。しかし、なぜ、それに疑問が生じないかいうと、こちらの方は事前に遺伝子解析がなされていませんから、どんな結果でも好きなように言えるわけです。「調査委が出した結果がこうなんだから、こうなんだ」と言われてしまえば、それまでです。しかし、B6由来のFI幹細胞の方は事情が違います。B6由来のFI幹細胞は、遠藤氏によってすでに遺伝子が解析されていますから、それと一致するサンプルとすり替えなければなりません。そんなことをするのは面倒だし、リスクも大きいので、「B6由来のFI幹細胞は見つからなかった」ことにしたのです。そういうことです。
13. 一言居士
2020年01月14日 13:02
2015年1月2日
調査委の報告が遠藤氏論文と食い違う件。
「遠藤氏の解析は正しいのか、捏造か、どっちだい?」という質問を受けました。正直、私も一瞬混乱しました。厳密に言えば、生データを知っているのは遠藤氏の方ですから、究極的には、私が遠藤氏論文の真偽を判定することはできない。しかし、少なくとも言えるのは、調査報告が遠藤氏論文と一致しないのは、おかしいということです。双方が一致しないということは、少なくとも、どちらか一方がウソを言っていることになります。もちろん、双方がウソという可能性もあります。
いずれにしても、遠藤氏は、「FI幹細胞はES細胞とTS細胞の9対1の混ざり物」という結論を出しました。それなら、調査報告でも、それと同じ結論が出てもいいはずです。ところが、調査報告では、そういう結論を示していません。これは変だと気づくべきです。
14. 一言居士
2020年01月14日 13:03
ですけれども、調査委の側にも言い逃れをする余地があります。‥‥「遠藤氏が解析したデータはB6由来のFI幹細胞であり、当委員会が解析したのは129B6由来のFI幹細胞である。当委員会が解析したFI幹細胞にはTS細胞の混入は見られなかったのである」‥‥と主張できるわけです。
しかし、仮に、遠藤氏の解析したB6由来のFI幹細胞を犯人がでっち上げるとすれば、どうすればいいでしょうか? 若山研にあるB6のES細胞を持ってきて、丹羽氏のTS細胞と「9対1」の割合で混ぜる必要があります。しかし、そうやって、サンプルをでっち上げても、その遺伝子の解析結果が遠藤氏論文と一致するという保証はありません。たぶん、一致しないでしょう。となれば、非常に話がややこしくなります。ですから、そういう事態を避けるために、「B6由来のFI幹細胞は見つからなかった」ことにしたのです。これが真相だと思います。
15. 一言居士
2020年01月14日 13:06
2015年1月2日
調査報告は完全なでっち上げである。
今回の調査報告は完全なでっち上げである。以下、その根拠をまとめておきます。
(1)FES1は、かつて若山研にいた人が作製したものである。その人は、小保方さんが若山研にやってくる以前に若山研を去っており、そのさいFES1をすべて持ち去っている。それが若山研に残されていたという証拠も全くない。したがって、小保方さんがFES1を盗むことは出来ない。
(2)したがって、小保方さんの冷蔵庫にあった129GFP・ESは、FES1ではない。
(3)にもかかわらず、「129GFP・ES=FES1」であるという調査委の遺伝子解析はおかしい。⇒これは致命的な矛盾です。
(4)129GFP・ESが若山研で作製されたものならば、若山氏がそれを「知らない」というのはおかしい。誰がそれを冷蔵庫に置いたのかを知っていて「知らない」と言っているのであれば、若山氏はその人物をかばっており、したがって、冷蔵庫にそれを置いたのは小保方さんではない。
16. 一言居士
2020年01月14日 13:07
(5)若山研に所属するES細胞のケースについて、若山氏が本当に「知らない」のであれば、なおさら小保方さんがそれを「知っている」はずはない。いずれにしても、129GFP・ESを冷蔵庫に置いたのは小保方さんではない。
(6)「FI幹細胞の培地でES細胞は全滅する」と丹羽氏が証言しているにもかかわらず、「FI幹細胞はES細胞が混入して増殖したものである」という調査委の報告はおかしい。
(7)少なくとも、FI幹細胞の培地に混入されたES細胞は遺伝子にダメージを受けているはずであり、それが混入以前のES細胞の遺伝子とピタリ一致するというのは、むしろ不自然である。
(8)遠藤高帆氏が「FI幹細胞はES細胞とTS細胞が9対1で混入したものである」と結論しているのに対して、調査委が単に「FI=ES」と断定したのはおかしい。少なくとも、どちらかがウソを言っている(両方ウソもありうる)。
(9)調査委が遠藤高帆の解析したB6由来のFI幹細胞を「見つからなかった」というのは、きわめて怪しい。おそらく、「遠藤氏の解析結果と矛盾しないサンプルをでっち上げることができなかった」というのが、その真相だと思われる。
以上です。
17. 一言居士
2020年01月14日 13:22
DORA氏は論文通りのSTAP細胞はあるとした。そして、GOF由来FI-SCはあるとしたのである。なぜならば遠藤氏が発見していると。しかし、遠藤氏はそのFI-SCの背景として登録されているのはF1だから捏造だと主張している。DORA氏は小保方さんが理研にデータ提出した後、記者会見発表を待ってから理研が正式にNCBIに登録したことに気づいていない。他者が関与している。それはともあれ、その上で、今Letter Extended Data Figure 5-c,dが真正な写真かどうかが問われている。最後の問いだと言ってよい。
戻ってきてくれないかなあ。
(DORA説からの発展)学ブログに対してLetter Extended Data Figure 5-c,dをザハリッヒに検討しないかと呼びかけましたが無視して相変わらずため息ブログとのじゃれあいです。両方とも照井本人のようですね。最近一研究者ブログが復活したという情報をTs.Marker さんブログで聞きつけたので、 すぐに一言居士の名前でコメントしましたが返事が無い。アク禁になってたんだが外れていた。でも今は復活したかな。試してない。
今、Ooboeさんにとって 信用できる書き込み場所は根本さんだけかな。和モガさんは止めてるから書き込むと迷惑する。Ts.Markerさんは議論を余り好まない。多分理系の現役の方ではないか。余り正体を現したくないんでしょうね。多く語るといろいろと情報が出てしまうからね。DORAさんも現役臭い。出世の妨げになってはいけないね。根本さんは弁護士なんじゃないかな。仕事が忙しそうだ。Ooboe さんが書きこむならあそこでしょうね。あそこはとても客観的な場所だ。類は友を呼ぶでしょうね。人はOoboe さんの居るところに集まっているんで学さんのところに集まっているわけではない。
一番いいのはパートナー氏がブログ開設することですね。楠本さんに拡散してもらったら普通の人がたくさん集まる。あの西岡さんのガンバレブログに今アップされている動画は12月の作製だが10万人以上が見てる。関心は消えてないのが分かる。ただし、あの動画の内容はあまり正確ではないね。
DORA氏は今は処世上の理由でSTAP細胞事件には関心を失っている模様。でもかつての分析は誰よりも正確ですね。和モガさんとTs.Marker さんは、アルイミオウジ氏も含めて、自分の興味で現象を追うところがあって、STAP細胞の不思議に関心がある。その方向だと、自分で小保方さんの研究を引き継いで実験した方がいいでしょうね。事件は別問題でもっと人間の行動動機を分析しないといけない。科学的専門知識だけではとても解けない。
その点、DORAさんは自分では解に至ったと判断してたんじゃないかな。DORA説には考え抜けはあっても、主張間に矛盾が無いんです。そういう風に全体を眺めて解こうとしているからね。
でも、DORA説だとなぜ若山さんが論文を取り下げたのかが説明できません。DORAさんは若山さんが嘘をついているのだと判断しているんです。小保方さんは無実だ。でも、若山さんがまさかESコンタミなんてさせるわけがない。じゃあ、なぜキメラができたのだと考えると、論文通りにできてたからだよと結論する。そこで安心してこの問題から降りた。ここまでとてもクリアで説明されてないことはあるが、説明されていることで矛盾していることはない。
説明されてないことの最大の疑問が、できてたのに、しかもアクセプトまでされてるのに、どうして若山さんが嘘をついてまでできている真正の論文を取り下げたのかということです。これは科学知識の範疇内の問題ではない。専門の問題ではなく、一般的良識の問題です。大学は学生に常識を教えるところだと諭吉が言った意味での良識ですね。
何か間違えたから取り下げたのだ。そうでなくちゃんとできてるのに一人大騒ぎして取り下げるはずはない。小保方さんに騙されていたということに気づいたから一人大騒ぎしたのだと、DORA氏が考えて居るかというと、そうではなく、私は犯人ではないと言ってるのは若山さんだけで、小保方さんも笹井さんも丹羽さんも誰も自分は捏造してないなんて言ってないという。そんなことを言うのは疑われている人だ。自分自身で自分が疑わしい人の言い訳じゃないかと。これは諭吉の言う良識で、専門知識は不要です。
小保方さんは手記を書いた。これが書かれずにそのまま姿を消していたら、小保方さんは犯人だと世間に思われたままだったでしょうね。若山さんは小保方さんをリクルートしようとしていた。このことは手記だけでなく、デイナ・グッドイヤーのヴァカンティへの聞き取りでも明らかになった。分からなかった動機の一つが世間の目に明らかになった。このことを言ってるのは手記だけで、桃子も聞いてない、NHK、日経サイエンスも知らない。デイナは手記を日本にいる友達に訳してもらって小保方さんにインタヴューした。やっぱりきっかけは手記です。
話は東北大震災の日に遡るのだ。小保方さんは震災の所為で若山研に腰かけたが、ビザが取れた時に引き留められて自分のラボに来ないかと誘われたが、ヴァカンティが反対した。小保方さんの希望を入れてGOFマウスでの実験を出来るように日本出張扱いにしてくれた。この辺りがとても妙で、若山さんはなぜ断らなかったかという問題がある。
推測ではこの時既に山梨大への転勤は決まっていたので、やはり助手で誘えば来てくれるのではないかと考えていたのではないか。というのも、帰国前に誘った時に自分のラボに来いと言っている。CDBでの10年の期限が迫っていているのに、その後どうしてやるつもりだったのかという問題になる。せっかくハーヴァードに就職できているのにそれを遮るのならもっと後まで責任を負わねばならない。だから、そういう推測が出来るということになる。すべては手記が書かれたから推測可能になった。
野依理事長がキメラができなかったとき、どうして返さなかったのかと疑義したが、そういう裏事情は桂報告書時点で誰もしらなかったのである。知っていたのは西川さんと若山さんと小保方さんだけだ。小保方さんが手記で語ったから、背景が理解できるようになったのである。NHKも桃子も日経サイエンスも知らなかった。何にも調査取材できてない。
若山さんは最初小保方さんの熱心さを気に入って山梨大に助手として連れて行きたかったという動機を持っているんですね。
この時、まだ何もできてないので小保方細胞に関心があったわけでは無いことが分かる。赤ちゃんマウスを使うように指導したころからGFPがよく光り出した。そして、ライブセルイメージング実験が行われていて、最初光らないものが徐々に光り出す。自家蛍光でないことは最初から分かっていて、後の丹羽さんの検証で、GFPの漏れ出しが発見された。
知られざるアーティファクトですね。この頃誰も知りませんから多能性指標が出たと信じた。若山さんも同じです。
これが知られざるアーティファクトであったのか、本当の多能性マーカー発現であったのかは、丹羽さんの実験が正しいのかどうか、論文通りの手法ではないのではないか等も問われますね。論文では内在性Oct4発現もちゃんと定量PCRで確かめられていてES並に出ているが、キメラが出来ているからということで結果を盛っていないかという疑義も出ているところです。でもArticle Figure 2-aの免染結果はスゴイですよね。捏造写真ならES細胞でしょうけど、彼女はメチル化実験でESを使わなかったから結果が出ないので、いろいろと操作したのだったはずですよね。ここでESならメチル化でも使うでしょ。でも使ってないのみならず、最初は正直に持っていったら、是では使えないと若山さんに言われている。
ともあれ、これだけ光っていてもキメラは出来なかった。論文は後から書かれたので、データは後から揃えられた分もあるでしょうね。小保方さんは図は論文論旨が理解されやすいように添付するものだと思っている。とても危険な思い込みでしょうね。捏造意図が無くても、事実を思いの方向に捻じ曲げる危険性がある。キメラが出来てないときはいろいろと事実に従って論旨を考えてますが、キメラが出来てしまうと、できたという事実に添って他のデータをあるべき姿に捻じ曲げてしまう。こういうのはどんな分野でも素人の陥りやすい陥穽ですよね。事実が論旨を作らないといけないという根本が飲み込まれていたら、キメラの成功と他のデータが矛盾していたら、真実により近づけるからうれしいのでなければならない。その矛盾を考えることによって自然と人間の思い込みとの違いに気づけて新発見がある。
キメラは出来ているのに脱メチル化はしていなかった。これはとことん解明しなければならない矛盾です。誇れるデータではないと自覚しているなんて問題ではない。自然科学者としての使命の自覚の問題です。何をやってるんだということですね。自然の真実を解明したいんじゃないのか。
これを指摘する側の精神構造もおかしいですよね。目くそ鼻くそじゃないんですか。白線を入れるルールなんてどうでもいいでしょ。データをいじっちゃいけないという規則の問題ではないでしょ。脱メチル化していないという事実はキメラがほんとには出来てない筈だという証拠なんでしょ。
小保方さんは自分で気づけないといけません。そういう姿勢だと科学者にはならない方がいい。
犯人は若山さんだと分かってしまったから言うのですが、小保方さんは別の分野で処世して行った方がいい。どんなに才能の有りそうな人を選りすぐっても、そもそも科学的新発見をする人は1000人に3人なんです。早く自分の才能が無いことに気づいたのは不幸中の幸いだと思わないといけない。997人の中に入ってくすんでいるより、別の分野で活躍したがいい。一番いいのは取り敢えず普通の主婦になることだ。これだと少なくとも大きくは人生は間違わない。保険掛けとく。ホーキング博士は自分の説が間違ってる方に友達と賭けたと書いてる。悲しさは勝ち金でコンペンセイトされる。かな? 神がお決めになっていたら、主婦になっていても又違う道が開けるでしょう。キューリー夫人は主婦ですよね。何事も神の御心のままに。
では翻ってなぜキメラができたのか。キメラができていたのなら、その時のSTAP細胞は大半が脱メチル化していたはずだ。若山さんが使った細胞は小保方さんの脱メチル化してない細胞ではなかったのか。
小保方さんは正直に脱メチル化してませんと先生のところに持っていった。それを若山さんはなぜどうしてかなと問わなかったのか。それでは使えないって何だ。
ここはキメラが出来ているのだから脱メチル化しているはずだ。あなたの検査テクニックの問題じゃないのかと押し返すのが自然でしょう。あなたちゃんとメチル化検証できるのかと。
ところがここに妙な事実がある。桂報告書ではこのメチル化実験は2011年10月27日 1 セット、同11月17 日に 2 セットGRASに依頼されたという。
キメラ成功前である。キメラ成功は2011/11/28である。>>
その後、2012年4月12 日付けの PR 資料に全く別の実験結果が図として提示され、 この図は2012年4月のNature投稿原稿、Cell投稿原稿でも使われ、最終的にArticle Fig.2cとして発表された。この図では、メチル化を示す黒丸の配置に一部乱れがあ り、手動で作図したと考えられた。また、CD45 陽性細胞(CD45+および Cultured CD45+) のデータについては、両者のパターンが酷似していた。
これでは使えないと若山さんが言ったから2012年4月12 日付けの PR 資料が捏造された。使えないと言われた元の資料は2011年10月27日 1 セット、同11月17 日に 2 セットのままだったから、使えないと言われた。
若山さんは脱メチル化していない小保方細胞を使ってキメラを造れはしないであろう。
2011/11/28に帝王切開された4 Nキメラは10日ほど前の11/17頃に小保方さんから渡されたことになる。小保方さんは2011/11/17のその日に第二回目のGRAS提出をしている。その時のSTAPですね。ES細胞は入ってませんね。ES細胞が入っていたら脱メチル化してたでしょう。ES捏造しようとしている人が、GRASにはそのまま脱メチル化してない細胞を渡すなんてことはありませんね。小保方さんは素直に作って、それを若山さんとGRASに渡したんですよ。一方はキメラができ、他方は脱メチル化してない真正な実験結果だった。若山さんはそれを見せた小保方さんに、それじゃあ使えないよと言った。
若山さんが何をしたから脱メチル化してない細胞からキメラが作れたのでしょうかね。ntES化したからだ。若山さんは脱メチル化程度が低いからキメラが出来ないのだと知っている。でもntES化することによってキメラはできる。そして大量にGFP蛍光している小保方細胞の性質をキメラに何が加わったかによって解明できると考えた。
- 2020/01/12(日) 18:23:56|
- AC129
-
-
| コメント:4































