d. TCR検証との関係(西川氏のアドヴァイス)AC129の実験が「遺伝的背景が及ぼす影響」を調べるためでなく、細胞追跡の実験だったらどうでしょうか。その可能性も念頭に置いて検討してみましょう。そうでないと、なぜこの実験に関して129ローザなんて話が紛れ込んでくるのかが分からない。何かまだ知られていないことがあるからとんでもない"勘違い"だという説明になってしまう。誰がローザと聞きもしないでローザと書くでしょう。誰かがローザと言ったからローザと書かれている。それを追求しないと、単に勘違いで済む問題ではありません。自分に置き換えてみたらわかる。聞きもしないで、かつ何の理由もなく、突然ローザと書けるか。はは。
我々の仮説では、若山さんは小保方さんの細胞がキメラ形成しないことは知っています。しかし、何物かではあると信じて、或は何物であろうかということを知ろうとして、小保方酸浴細胞核を使用してそのntESからキメラを作ってみている。そしてどうもその胎盤が光っているのではないかと気づいたわけです。だからこそそのntESをTS培地によって誘導してCTSを作ろうとした。
ただし、この胎盤蛍光が単なる希望的思い込みでなかったかということが後の議論でも出て、かつ小保方さんの免染も確かにGFPは入っているんですが正しくその場所を免染しているとは限らない。つまり光って当たり前の場所を調べてたのではどうしようもないという疑義がでたわけです。このことは笹井さんが参加してからのことで、査読者たちに指摘されたり、事件発覚後にパブピアで指摘されたことなので、この当時は論文も幹細胞化の実験に関しては発表していないから、胎盤の件が正しいか否かは二人ともまだはっきりとは分かってないんです。
一方でACCsの論文がネイチャーにリジェクトされ、若山さんにしてみたらリクルート絡みの問題は終わりだと思ってたら、ヴァカンティが高いハードルを飛べと小保方さんに指示したもので、論文はティシュー誌でなくセルとサイエンスに投稿することになった。
どうしたら通るかという小保方さんの相談を聞いてもらうために若山さんが西川さんのところに連れて行った結果、TCRアドヴァイスを得た。そして実験して結果を添付したがまたセル、サイエンスともにリジェクトされた。
(PCRの結果)一方でFI幹細胞の研究をつづけながら、若山さんは小保方さんからのTCR結果を聞かされている。彼は小保方さんの作ったリンパ球の酸浴細胞にTCR再構成PCR結果が出るのは当然だと思ってる。CD45+だけでも出るものを念を入れてFACS選別で確率高くT細胞だけに絞ってます。ある程度違う細胞が混じってもPCR検査では出るに決まっているし、現に出てる。もう1度貼り付けましょう。
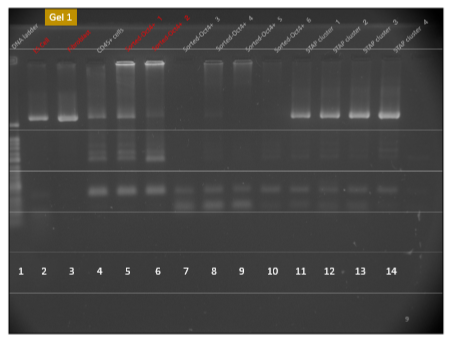
ど素人なんでどれがTCR再構成バンドか分からなかったから苦労しましたが以下が泥縄のお勉強の成果でした。
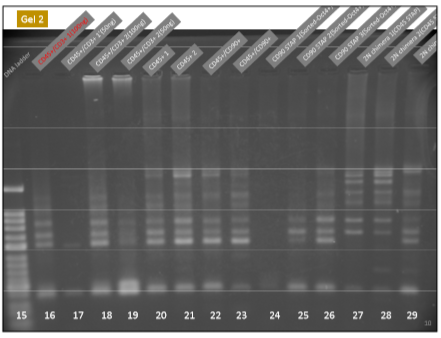
はっきり出てますね。CD45陽性細胞と酸浴STAP細胞にはTCR再構成はあります。ここは問題ない。キメラにもTCR再構成があるが、ここはどういう出方をしているかが問題になった。そしてGel1とGel2には所謂STAP幹細胞のPCR結果がありませんが、幹細胞は別のラボメンバーが担当した。これが後に問題になった。
ここで残されている課題は
キメラと幹細胞にはどういうバンドが出ているべきなのかという問題です。
STAP幹細胞は若山さんが小保方さんから渡された細胞を基本ES培地で培養誘導して作られたと若山さんが語っているものを小保方さんが論文にプロトコルとして書いている。手記によれば小保方さんは自分ではプロトコルに従っては作れていない。作れないのに構わずに書かせたのは若山さんですが、丹羽さんも相談を受けて若山さんを信じるべきだと答えている。こんな論文はおかしいと誰でも思う。でも、笹井さんと丹羽さんは業務命令でリヴァイズの手伝いをしていますからね。実験が疑わしいというスタンスは初めからありません。
筆頭著者ができないのに責任著者が自分はできるから、できたと書いて置けと言われ、シニアの指導者も若山さんを信じるべきだと助言した。後から、若山さんはあれは小保方さんがESを自分に渡したのだと言い出した。丹羽さんは梯子を外されたんやと、その足で山梨に向かったという。記者会見でなにしろESを渡されたと思い込んでいるもんでと、語っている。
ESを渡されたら誰でも気づくということは既に我々が証明した。若山さんは嘘をついている。論文は通らないようにと書かせているんですね。それが通ってしまったんで、逃げようとしたんです。通りそうだと寺下さんから情報を受けた頃からあちこちに情報リークをして準備を始めている。そしてタレコミがあって事件化するまでは知らぬ顔をして小保方論文をほめていた。11次元の博論のテラトーマ画像流用指摘から突如豹変した。予定通りです。11次元に博論の件をあらかじめ知らせたのは若山さんですね。
ここから先はかなり悪質ですね。どうしてこんなことになったか。何かやはりリクルートの経緯だけでなく、若山さんを心情的に怒らせるものが加わっているのではないかとも疑義される。事件に無関係なはずの遠藤氏の、舐めてますねこれという、やくざ的な言葉づかい、或は真綿でじわじわなんていう、会ったことも無い相手に対するいわれのない憎しみの原因がどこにあるかと推測すると、若山さんからの情報ですよね。俺達を舐めてると。どういう情報のやり取りになっているのでしょうかね。
(研究の発案と特許)若山さんは小保方さんが山梨に来てくれると思って、或は来て欲しいと強く望んで、幹細胞化の論文のためのデータを渡している。ところが小保方さんはそのデータを持ったままヴァカンティの許に帰った。
Ooboe さんとパートナー氏の公開請求で共同研究契約書は交わされていないと判明している。理研の事務窓口は共同研究の実態があったのかどうかも分からないと答えている。
すると小保方さんの客員身分はどういう手続きで生じたのか。どうやら、客員受け入れ申請書の類のものらしい。そしてその保管期限も過ぎて書類は無いという。
では客員申請はどういうケースで行われるかというと共同研究申請書に基づくのが普通であるが、今、理研のその規定は見れなくなっている。どうやら安易なイレギュラーな客員受け入れが行われていたようですね。そしてそういう受け入れは海外でもあって、学者が海外旅行のために休暇を取るのにこの契約がまま流用されているらしい。このケースでは小保方さんの震災避難の受け入れ、或はそれ以前の博論実験の受け入れでもそのイレギュラーな処理が行われているのではないかと疑われる。契約書が無いというのは正式な理研の答えで、かつ、岸氏がちゃんと約定してないからこうなると怒ったという情報も流されている。
そういう研究状況の中で、小保方さんの論文の投稿と、若山さんの幹細胞化の実験が並行して進められている。スフィア細胞自体はヴァカンティ研での小保方さんの発見であるが、理研に来て酸浴実験からGOFの赤ちゃんマウスを使ってのOct4-GFPの蛍光までの実験は若山研での発見である。しかし、約定の無いままになんだか分からない発見らしきものが次々に続いていく。
特許でもヴァカンティと揉めている。11月に小保方さんがヴァカンティの許に戻った時、幹細胞化実験のデータは小保方さんが持ち逃げした形になっているが、渡したのは若山さんで、しかも共同研究契約書は存在してない。小保方さんが若山さんに渡されたデータは一体誰のものなのか。
岸氏が約定もなくこんなことをしているからこんな問題になると怒ったらしいのは一般人でも当たり前だと思うところでしょうね。でも本当は若山さんはそのままヴァカンティ研で小保方さんが研究を続けてくれていた方がありがたかったでしょう。何しろntES化しないでキメラの再現はないし、幹細胞化の実験の意味も分からないはずだ。どうしてあの時キメラができたのか。別の実験のESをコンタミさせてしまっていたみたいと言い訳したらそれで済む。
ところが小保方さんは理研に呼び戻されてしまった。しかも笹井さんと丹羽さんがメンターになるという。とうとう本当のことを言えないままに論文が自分の手を離れてしまった。笹井さんの手にかかったら論文は通るかもしれない。キメラはスタンダードな方法でできたと書かれる。再現ができないと最後に自分のところに責任が来る。論文は通ってもらいたくない。せっかく三誌は落としてくれたのに今更別の雑誌に又投稿するなんて困る。それには幹細胞化の論文をつけさせて、一度落としてくれたネイチャーに再投稿するのがいい。こんなntES化実験をSTAP細胞からダイレクトにできたなんて書いて通るはずがない。小保方さんを山梨に連れてくるために言った勧誘言辞をそのまま利用して、そんなこととはつゆ知らない無関係な笹井さんに押し付けたことになる。山梨に来てくれたらこの幹細胞化の論文は小保方酸浴細胞の核を使用したntES化実験だと本当の研究論文を書かせることができる。でも、それを知らないままに笹井さんが論文を書いたら、いくら何でも通るはずは無いと考えたんですね。それで強硬にネイチャーへの二報同時報告に拘った。例えばPNASに笹井さんの論文構成力と信用力で投稿したら通るでしょう。そしてキメラができたということに焦点が当たる。そこは自分の責任のあるところとなる。困るんですね。だから何が何だか分からなくする意味で幹細胞化論文の二報同時投稿を強硬に主張したんですね。
ここでどうして本当のことを言わなかったのでしょうかね。やっぱり人事のことでいろいろと自分で売り込んでるんでしょうね。特に西川さんがメンターですからね。今まで話してきたことと矛盾するようなことがあると話せないですね。論文が通らなければそれが一番無事なんですね。
小保方さんに幹細胞が自分ではできないと訴えられても、作り方を目の前で教えもしていない。論文は通ったら困るんですね。こんな論文はトンデモなんだから通るはずはない。通るように教えるなんて輪をかけてトンデモだ。僕の手技だよとごまかしてシカトしている。そんな科学は普通ではあり得ません。
若山さんの誤算は自分はntES化させたことを知っていて、ntES化させないでキメラができるわけが無いと分かっている。査読者たちは誰も信じないと思い込んだことです。現に三誌でリジェクトされている。でも、笹井さんはそんなことは知らない。ここで起きていることはすべてダイレクトに小保方細胞が引き起こしている現象だと信じて論文を構成している。
若山さんが通るなよ、通るなよと念じているのに、笹井さんは通るように通るように努力した。理研と笹井さんと丹羽さんの信用力が加わって、理研と若山さんだけの信用では通らなかった論文が通りそうになってしまった。
小保方ユニットリーダーの元には寺下さんがついていて、若山研とは常時連絡を取り合っている。若山さんは追い詰められてしまったんですね。
(出るべきバンド)幹細胞とキメラ体細胞からはどういうTCR再構成バンドパターンが出るべきなのか。CD45と酸浴STAP細胞からは全部のパターンが出る。こちらは簡単で、現に出ていますね。T細胞を選別しているのですから当たり前です。
でも、幹細胞とキメラは違いますね。まず幹細胞の方は、小保方さんの酸浴STAP細胞を基本ES培地で培養したら増殖を始めたということが本当なら、これはSTAP細胞と同じく全バンドが出ます。すべての幹細胞株でTCR再構成の全バンドがでる。それは細胞集団がポリクローナルな集団だからです。
ところが、この実験を若山さんは小保方さんにさせなかった。これを確認したのは別のラボメンバーで、結果は調べられた8株にのうち2株にはTCR再構成があると、小保方さんは知らされているように手記の149Pに書いている。ラボの中での報告でも、それを合わせて報告していると思われるが、桃子本ではあったと本人が報告したと書かれ、桂報告書でも相変わらずあたかも小保方さんが全部行ったかの如くミスリーディングな書き方をしている。
8株のうちの2株にあるという意味はとても不可解なものです。論文のプロトコル通りに作られているものなら、すべての株に全バンドが出ないとおかしい。
一個一個の細胞のTCR再構成のありかたの可能性の全ては以下ですね。我々のお勉強の成果でした。もう一度確認しておきましょう。小保方さんのプライマーで挟むとカッコ内の6本のバンドが出る。7~12は検出できないということです。J1、J2ともセグメント数を6としています。実際には7つのようですが、再構成するときの仕組みが素人には理解がちょっと難しい。でもそれはこの問題にはあまり関係しない。
>>
01.D1-J1の123456-D2-J2の123456(*GL)
02.D1-J1の23456-D2-J2の123456(*GL)
03.D1-J1の3456-D2-J2の123456(*GL)
04.D1-J1の456-D2-J2の123456(*GL)
05.D1-J1の56-D2-J2の123456(*GL)
06.D1-J1の6-D2-J2の123456(*GL)
07.D1-J2の123456
08.D1-J2の23456
09.D1-J2の3456
10.D1-J2の456
11.D1-J2の56
12.D1-J2の6
13.D1-J1の123456-D2-J2の234567(*リアレンジバンド)
14.D1-J1の123456-D2-J2の3456(*リアレンジバンド)
15.D1-J1の123456-D2-J2の456(*リアレンジバンド)
16.D1-J1の123456-D2-J2の56(*リアレンジバンド)
17.D1-J1の123456-D2-J2の6(*リアレンジバンド)
ポリクローナルな細胞集団をPCRの検体にすると全部のバンドが出るということです。論文通りの幹細胞の作り方だとそうならないとおかしい。8株の中に2株ほど"あったようだ"というような出方にはならない。
しかし、検体がモノクローナルな細胞集団であると、この中の二つの組み合わせになる。というのは染色体が二本あるからです。従って、上記のリストの二つの組み合わせになる。結果的に0本か、1本か、2本ということです。
でも、小保方さんも、手伝ったラボメンバーも8株の検体がモノクローナルな細胞集団だなんて予期していませんよね。丹羽さんは後にこれをもう一度検査させて、おかしいと気付き論文から外すように笹井さんにアドヴァイズしました。流石ですよね。ただし、ntES化されているなんて考えもしてませんね。何か予期しない実験上の手違いがあると考えて後でゆっくりやり直したらいいと考えている。捏造なんて疑ったら手伝い自体をつづけられませんね。
キメラの場合はもっと複雑ですね。今度は小保方細胞そのものであっても、小保方細胞核使用ntESであっても、20個程度移植した個々の細胞の増殖していった器官部分は0本か、1本か、2本のいずれかのバンドしか出ない。狭い範囲を試料採取するとそうなるが、広い範囲で採取するとポリクローナルな結果と同様になる可能性もある。キメラの場合の尻尾の細胞がモノクローナルに構成されているのか、ポリクローナルに構成されているのか、事前に誰か研究でもしているのか。まして、2Nの場合はリシピエントの細胞も考えないといけない上に、組織の採取時点で、リシピエントからの白血球が混じってないという可能性も排除されているかどうかが書かれていない。方法論自体が厳密さを欠いている。丹羽さんと笹井さんが幹細胞とキメラのTCR再構成結果を外させたのは当然ですね。ただ、キメラはスタンダードなやり方でできているということは疑ってないから、捏造に気づくなんてことは誰にもできません。キメラは出来たと聞いていて、それが前提で論文の書き方を指導してやってくれと頼まれているだけだ。捏造だと思ったら途中で降りるでしょ。そして捏造調査に入るでしょ。
11jigen氏が笹井さんの立場に居たら、同時に国会図書館に行き、博論を調べ、ティシュー論文も調べて突合せるということを、論文のリヴァイズを行う傍らで3か月間で両方やれたというのでしょうか。そんなことを疑うのだったらこの仕事は引き受けないでしょう。そもそも11jigen氏が博論を国会図書館にまで調べに行って、コピーまで取ってきたのは自分の自発的行動なのか、それとも誰かに情報を齎されてのことでしょうか。
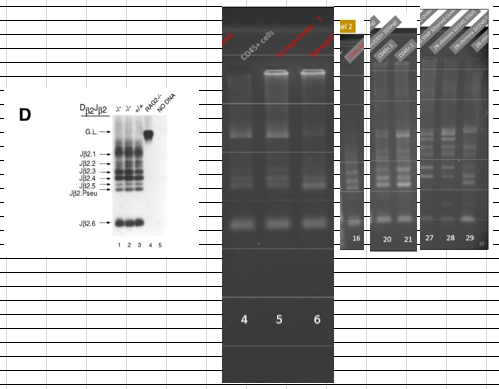
(オスしか居ないことを不思議に思わない不思議)2NキメラのTCR再構成PCR結果がGel2と特許申請書に存在している。これがとても錯綜した結果です。Gel2の2Nキメラです。一本欠如がある。難問です。
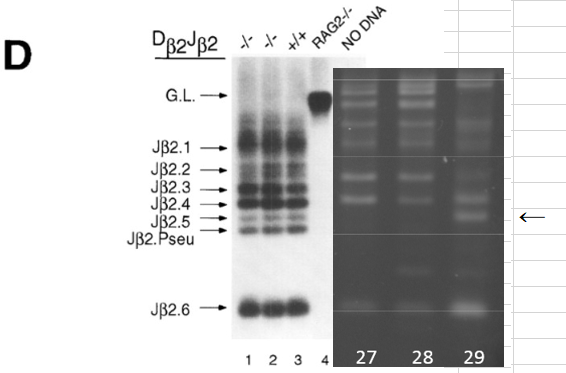
取り敢えず幹細胞の雌雄の問題から検討してみましょう。桂報告書の検査結果は以下です。
>>
①FLS1~8 オス
②CTS-1,11~1 オス
③GLS-1~13 メス
④AC129-1,2 オス
⑤FLS-T1,T2 オス
⑥GOF-ES メス
⑦129B6F1ES1 オス
⑧129/GFP ES オス
⑨FES1 オス
⑩FES2 オス
⑪ntESG1 オス
⑫ntESG2 オス
まず、FLSは現状は全部オスです。Article Extended Data Figure 8-jがそのFLS8ラインの4Nキメラのジャームライントランスミッション実験の結果です。
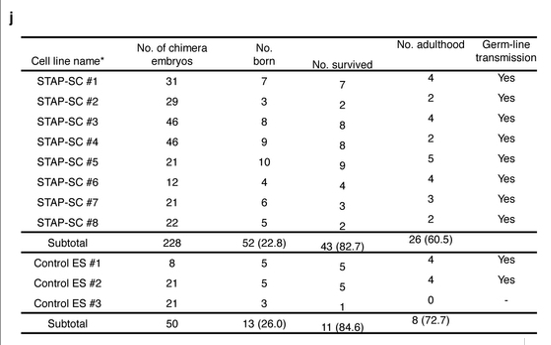
以下がそのリジェンドです。
>>
j, Production of mouse chimaeras from STAP stem-cell lines by the tetraploid complementation method. *These STAP stem-cell lines were generated from independent STAP cell clusters. 4Nキメラということは胎児の体の組織すべてがオスだということです。従って精原細胞もオスです。要するにオスのマウスということです。
8ライン由来4Nキメラは全部雄です。従って兄妹交配は出来ません。相手のメスにGFPの無いものを持ってこないとジャームライントランスミッション確認ができない。「僕のマウス」のGFPがどうこう言う以前に、この時点ですでに若山さんがどうしてオスだけなのかということを不思議に思わないこと自体が彼の嘘を証明しているんです。8ライン作って全部オスなんて、仮に正しく小保方さんのスフィア細胞を使って作っていても、或は我々が判断しているように、小保方さんの細胞の核を使ってntES化したにしても全部がオスだったなんて確率はほとんどありません。そうではなくて、一つの成功したntESがたまたまオスでそれを8つに株分けしただけか、クローン胚はたくさん作っているはずだから、成功しているntESラインは複数あってそれを持っているのだが、小保方さんにこれがFLSの8ラインだから無限継代が可能かどうかちゃんと確認しなさいと渡していた8ラインが成功した複数のntES株の中の一つだけを8株に分割しただけのものであるかのどちらかでしょう。
既に、桂報告書が調べたFLSが何であるかは明らかになっています。小保方さんが普段使っていたFLSです。


松崎氏が2014/10/21に持ち出して調べたFLSサンプルは木星リスト123番なんです。13というのは上のGLSの13です。小保方さんが間違えてこちらに入れてるんです。FLSは8ライン、GLSは13ラインです。このFLSの全ラインがオスだったということです。でも、ここで雌雄を調べたとは限りませんね。なぜならこの調査は若山さんが先回りして無断で行っていますよね。以下です。

FLSが8株全部オスだということは調べなくても若山さんは知っています。なぜならFLSキメラにメスがいなかったから129/SvのGFP無しの雌マウスと戻し交配してジャームライントランスミッション実験をしたんです。「STAP細胞は産仔を数匹混ぜて作るため、性別が一致する率は高くない」って、今更何言ってんでしょうか。ジャームラインの実験の時には分かってやってたんでしょうよ。
そして、この頃に生まれた子の半分にしかGFPが来なかったと小保方さんに言った。つまり、この実験が何かコンタミがあったかもしれないという言い訳をこの時に準備しているんです。それはこの段階ではもっぱら米国特許仮申請をしてしまったヴァカンティに対する言い訳です。
又、小保方さんがリヴァイズ中に丹羽さんとPCR再検証してTCR再構成が無かったとされた実験後のサンプルが、木星リストの2番、4~10番のP2とされている8ライン分ではないでしょうか。笹井さん丹羽さんはこのサンプルを世界に公開しようとしてプロトコルを書いたんですね。その時にこの幹細胞にはTCR再構成は無いと書いた。手記の説明はそうなってますね。キメラ実験を信じているんですから当然の処置ですね。
それをTCRがないのに捏造を疑わないとは何事だと騒いだ最初が遠藤氏でしたね。舐めてると書いてましたね。無関係な人が突然入り込んできて丹羽さんに対して舐めてるなんて学者とも思えないやくざめいた唐突な書き込みが、kahoの日記でした。この人が何者なのかは11jigen氏と同様いずれ検討されなければなりません。
ともあれ、我々のntES仮説であると一種類だけを株分けすると全株がオスだけということは当然あるのですが、先に述べた2Nキメラの尾部のTCRに欠如しているバンドがあって、これは何種類かのntESラインを混ぜている可能性しか考えられないところなんですね。リシピエントの血液が混じっていたら全バンド出ます。
キメラのバンド欠如を考慮すると、我々はたくさん作ったntESをいくつか混ぜてキメラ胚に挿入したのではないかと推測したのですが、
幹細胞に関しては1種類を8株に株分けしただけのような結果になっている。桂調査は太田ESのコンタミ捏造という線で論旨を構成しているのでntES説に関しての証拠が不足していて、なんとも分かりにくいところです。ただ、今我々は太田ESのコンタミは無いと証明してしまってますからね。ここは本物かntESしかありませんが、幹細胞がポリクローナルな集団で無かったことで既に、論文通りの幹細胞でないことも証明されてしまってますよね。幹細胞はモノクローナルな細胞株なんです。桂報告書27P。
>>
1)TCR 遺伝子再構成に関する不整合データ隠蔽の疑いについて
(調査結果) 小保方氏は TCR 遺伝子再構成に関する実験を開始し、STAP 細胞を含む細胞塊、一部の STAP 幹細胞に TCR 遺伝子の再構成が見られることを CDB 若山研で最初に報告した。しか し、後に 8 系統の STAP 幹細胞の TCR 遺伝子の再構成を確認したところ、再構成は確認 されなかった。なお、この8系統は小保方氏が継代培養を繰り返していた細胞であった。
さらに、この実験は小保方氏の依頼で、CDB 若山研メンバーによる TCR 遺伝子再構成 の確認実験が行なわれた。しかし、この CDB 若山研メンバーの実験ノートによれば、実 験の結果 TCR 遺伝子の再構成は確認されなかった。
以上のことから、小保方氏は最初の実験でTCR遺伝子再構成があることを報告したが、 後の小保方氏自身の実験、および CDB 若山研のメンバーに確認を依頼した実験では TCR 遺伝子の再構成を認めるに至らなかったことから、実験データに不整合が存在したこと は明らかである。
丹羽氏は 2013 年 1 月に論文作成に加わった際に、小保方氏が継代培養を繰り返して いた 8 系統の STAP 幹細胞の TCR 遺伝子の再構成は確認されなかったと聞いたと説明し ている。さらに、丹羽氏は笹井氏に対して、TCR 遺伝子再構成に関するデータを論文に 含めることについては慎重にすべきとの意見を伝えた。小保方氏の追試が不成功であっ た点に関して、笹井氏らは STAP 幹細胞がヘテロな集団であり、長期的な継代培養により再構成が起っていた細胞が消失したという解釈を採った。なお、Article 論文には、 STAP 細胞を含む細胞塊の TCR 遺伝子再構成については記載されたが、STAP 幹細胞自体 の TCR 遺伝子再構成実験の結果については記載されなかった。
一方、丹羽氏は、Protocol Exchange への投稿は、発表後、この論文ではすぐに再現 性についてクレームがつくと思った。小保方氏のプロトコールでは不十分と考えそれを 詳細にしたものを早急に公表すべきと考えた、と説明した。さらに、当時、小保方氏と 笹井氏はコリジェンダム(corrigendum)で相当に多忙であり、エディターと応答でき る者が必要ということで、自分が執筆した、と説明した。
2014 年 3 月 5 日に Protocol Exchange に公表された詳細なプロトコールの「STAP stem-cell conversion culture」「 2.After 4-7 days of…」のプロトコールの「IMPORTANT」 (iii)に、8 系統の STAP 幹細胞には TCR 遺伝子再構成が認められない、という結果の記 載が存在している。
また、丹羽氏は「若山さんは、最初 STAP 幹細胞の初期のパッセージでは TCR 遺伝子 再構成はあった、と小保方さんから聞いたと言っている」と説明した。
(評価)
TCR 遺伝子再構成に関しては、最初小保方氏が再構成を確認したとされたが、その後 の CDB 若山研メンバー、および小保方氏自身の追試で失敗した。その事実にもかかわら ず、実験結果を自分たちのアイデアに沿うようなものを採用したものの、後に、Protocol Exchange で 8 系統の STAP 幹細胞には TCR 遺伝子再構成が認められないという結果が記 載されたこと、並びに丹羽氏への聞き取り調査における上記の説明から、意図的な隠蔽 ではなく、研究不正とは認められない。 小保方さんの手記での説明と矛盾していますね。手記で書いている幹細胞サンプルを世界に公開するという目的があって、プロトコルにはTCR再構成が無いと書いたのだという事情を桂報告書は意図的に外していますね。そしてTCR再構成が無いということを捏造だと気付くべきことだと世間に間違った印象を与えるように文章を構成している。キメラが出来ているという大前提の下で幹細胞のTCR再構成実験に何らかの間違いがあることは、後にちゃんとやり直せばいいことで、キメラの捏造を疑うのならまず若山さんが何をしたかから調査しなければならない。しかし、彼らは若山さんの主張している通りに調査を始めた。犯人の指示に従って調査しているんですね。御笑い種です。警察を入れないからこんなことになる。
「さらに、この実験は小保方氏の依頼で、CDB 若山研メンバーによる TCR 遺伝子再構成 の確認実験が行なわれた。しかし、この CDB 若山研メンバーの実験ノートによれば、実 験の結果 TCR 遺伝子の再構成は確認されなかった。 」と書きながら、だから幹細胞にTCR再構成があるなんて書かれてないでしょと言うことに気づかないわけも無いと思われるが、もしここで、桂チームが丹羽さんたちが気づくべきだと思ったのなら、どうして若山さんが何をしたかを調べなかったのか。何を勝手に太田ESのコンタミだという若山さんの嘘に騙されているのか。若山さんが犯人だということに気づけば調査サンプルの中身が入れ替えられているということにも意識が向くでしょう。笹井さんたちがこの実験が捏造だと気付かなかったことをとやかく言う前に、お前たちがなぜ若山さんが犯人だということに気づかなかったかを先に反省すべきだと、警察なら言うでしょうね。プロは人を疑う時には先にすべてを公正な立場で検証する。第三者調査といいながら、若山さんが犯人だという可能性を最初から除外している。捜査のドシロウトと笑われてしかるべきでしょうね。尤もど素人だったからなのか、意図的な隠蔽だったのかはこれから暴かれていくでしょうね。
今、Ooboe さんのパートナー氏がその細胞の入れ替え問題に関して検察に申請書を提出された。ES細胞を小保方さんが若山さんに気づかれずに渡すことはできないと既に証明されている。にも拘わらずFES1はFLSと同じだという調査結果がでた。ブーメランですね。これがサンプル入れ替えの動かぬ証拠です。FES1の中身は若山さんが入れたFLSです。このことは和モガさんが既に近縁率の数値結果からも導いていますね。和モガブログで確認されてください。(細胞増殖率グラフ)FLSに関してついでですから調査されていないP40のことに触れて置きましょう。FLS1~8の細胞増殖率表は小保方さんが正しく3日ごとに40継代しているからP40のサンプルが残されているんです。もう一度掲載しましょう。

小保方さんはこの中の一つのデータを増殖率表に使用したんです。桂報告書が海外出張等で3日に一度出られた日はないと言ってるのは小保方さんがES細胞の増殖率データに関して、2011年の春と嘘をついたから、まだ腰かけているだけで客員にもなってないときですから当たり前ですが謝金支払表になかったんです。
ES細胞の増殖率なんてわかり切っていてわざわざ金をかけて新たにやらなくてもデータはどこにでもある。若山研の予算でそんな無駄なことを若山さんがやらせるわけがない。小保方さんは咄嗟のことで事情も分からないままに、若山さんを庇うための嘘をついた。桂報告はそれを承知の上で捏造指摘している。大事なのはSTAP細胞の増殖率とFLSの増殖率で、それはちゃんと行われていて、Ooboe さんのパートナー氏の取り寄せた謝金支払表ではほとんど出勤している。ついでですから証拠保全をあちこちに置くという意味でも楠本さんのデータをここにも貼り付けて置きましょう。

桂報告書はこの件に関して以下のように結論している。ESの増殖率実験が実際には行われていないことを、FLSが行われているにも関わらず意図的にレトリックを弄してあたかもどちらも行われていないかのごとくに書いている。
>>
(評価) この実験は行われた記録がなく、同氏の勤務の記録と照合して、Article Fig.5c のよ うに約 3 日ごとに測定が行われたとは認められない。"この実験"というのは正確にはES細胞の実験のことだけを言ってるんです。でも勤怠を見てない人はFLSも行なわれていないと思うでしょう。でも上のサンプルに小保方さんの字でP40と書かれたFLS-1~8があります。そして2月から5月にかけての120日間3日ごとに出勤できないような箇所はありません。そもそも腰かけていた2011年の春はまだ蛍光細胞すらできてなくて、こんな時期に分かり切ったESの増殖率実験を小保方さんがやってるわけがないし、まだ客員にもなってないのに勤怠があるわけもない。そんなことは桂チームも分かってるんです。小保方さんが何が起きたのか分からない状態で、とっさの嘘の言い訳をついたのを利用して間抜けなレトリックを弄して捏造虚偽報告しているのです。
(再度雌雄の問題)本題に戻って、今ある通りにFLSが全部オスだけだったらメスは生まれてこないんで兄妹交配はできないから別のジャームライン確認用の雌を持ってこないといけないと分かったところです。Article Extended Data Figure 8-j は4Nの実験だからそうなる。では2Nだったらどうなるのか。それがひとつ前の図のArticle Extended Data Figure 8-i です。
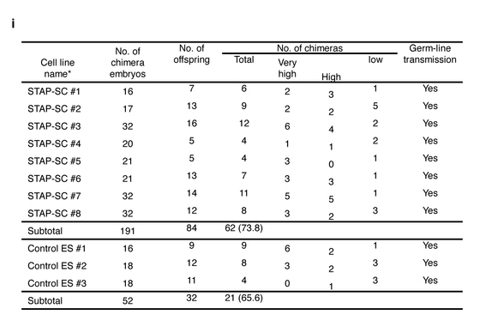
リジェンドは以下です。
>>
i, Production of chimaeric mice from STAP stem-cell lines using diploid embryos. *These STAP stem-cell lines were generated from independent STAP cell clusters.ここにはメスがあります。なぜならリシピエント卵のインナーセルマスから雌ができるからです。GFPがあろうとなかろうと兄妹交配してGFPが来たらジャームライントランスミッション有、つまりYesとなる。
アーティクル本文のマテメソに以下のようにある。
>>
Because the number of CD45+ cells from a neonatal spleen was small, we mixed spleen cells from male and female mice for STAP cell conversion. To make germline transmission more efficient, we intercrossed chimaeras in some experiments.ここで小保方さんが書いていて、かつ笹井さんが英文をみてやったintercrossの意味は兄妹交配(sibling mating)のことで、backclosss(戻し交配)と対照的に使っているのではないかと思われますが、正しい使い方か否かは素人なのでよくわかりません。近交系マウスを作るときにはincross,cross,backcross,intercrossという概念があって、それぞれ、雌雄相互に同一遺伝子をホモにもつ個体同士の掛け合わせ(完成した近交系マウスの掛け合わせはすべての遺伝子座でインクロスになる)、雌雄相互に異なった遺伝子をホモに持つ個体同士の掛け合わせ、ホモとヘテロの掛け合わせ、ヘテロ同士の掛け合わせという概念です。
この定義に従うと、STAP細胞の場合、そもそもF1マウスを使っている段階から先はできた子供同士の掛け合わせやワイルドタイプとの掛け合わせは全部インタークロスになります。そして別の近交系マウスを持ってきて掛け合わせたときだけバッククロスしていることになる。用語として正しいのかどうかはわかりませんが、他方メイティングの実際は論文で出ているわけです。
以下はArticle Extended Data Figure 7-bと7-cのSTAPキメラとそのジャームライントランスミッション実験の表です。
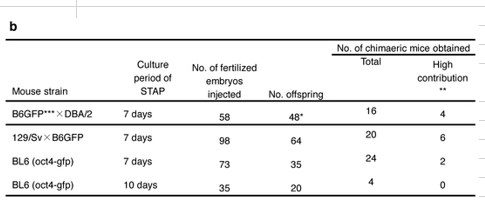

リジェンドは以下です。
>>
b, Generation of chimaeric mice from STAP cells by cluster injection. STAP cells used in the experiments above were generated from CD45+ lymphocytes of multiple neonatal spleens (male and female tissues were mixed). *All fetuses were collected at 13.5 d.p.c. to 15.5 d.p.c. and the contribution rate of STAP cells into each organ was examined by FACS. **The contribution of STAP cells into each chimaera was scored as high (>50% of the coat colour of GFP expression). ***B6GFP: C57BL/6 mouse carrying cag-gfp.
c, Production of offspring from STAP cells via germline transmission. Chimaeras generated with 129/Sv × B6GFP STAP cells (obtained from the experiments shown in b) were used for germline transmission study. b図は2012年初頭の実験です。DBA/2とのF1も使われている。この実験は2Nキメラ実験です。リジェンドのaにも書かれているんですが、右端にドナー貢献度が書かれているからそれだけでも判断できます。4Nキメラであれば胎児は全部ドナーですからこの記載は不要です。
そしてこのb図の2段目の129/Sv x B6GFPのキメラのジャームライントランスミッション実験がc図です。b図でキメラになっているのが20匹(内 High contributionが6匹)です。
2Nキメラですので何を根拠にキメラになっていることを調べたかというと、注に**The contribution of STAP cells into each chimaera was scored as high (>50% of the coat colour of GFP expression).とあって、GFP蛍光している毛色の面積が50%超をHigh contributionとしているので、その他も体毛のGFP蛍光判断です。そして紛らわしいですが、*All fetuses were collected at 13.5 d.p.c. to 15.5 d.p.c. and the contribution rate of STAP cells into each organ was examined by FACS.と書かれているのはあくまでも、最上段のB6GFP x DBA/2の子供だけで、これは帝王切開で全部取り出して、各器官組織を取り出して解剖し、FACSでGFP貢献度の確認をしたということです。従って二段目の129/Sv x B6GFPは出産されたマウスで、GFP蛍光の確認は体毛だけです。
この20匹から9匹を選び、ワイルドタイプを1匹加えてメイティングさせた。それがc図です。
2Nキメラですから体毛が蛍光しているからと言って生殖細胞までドナーだとは限りません。体毛判断でHigh,Medium,Low と分けているんですが、何か意味があるのかどうか分かりません。精原細胞、卵原細胞というのはとても早い段階で偶然の位置関係で決まるのではないですかね。毛色の貢献度とどういう関係なのか、よくわかりません。
結果、105匹の子供が生れて体毛のGFPもしくは黒目判断(リシピエントのICRはアルビノなので赤目)でジャームライントランスミッションの確認されたものが35匹ということです。これは見えるところにありさえすればいいので、見えないところで(例えば心臓で)ひょっとしたらあるかもしれないですが、実験全体としては一つでもあればトランスミッション有ということなので他は関係ない。三段目の実験は32匹出産されて体毛と赤目判断では0でしたが、それは心臓にはGFP蛍光があったかもしれないが、他で既にトランスミッション確認されてますからSTAP細胞はキメラ形成能だけでなく、更にジャームライントランスミッション能もあるという証明終わりだということです。
幹細胞キメラは別ですが、このSTAPキメラに関しては、すでにTCR再構成のPCR結果がサイエンスで書かれていることは査読文書で分かります。
>>
The DNA analysis of the chimeric mice is the only piece of data that does not fit with the contamination theory. But the DNA fragments in the chimeras don’t look the same as those in the lymphocytes. This assay is not properly explained. アーティクル論文とは違っているようですが、キメラのTCR結果も入っていますね。これらを丹羽さんと笹井さんが外させたんですね。
(B6の3番染色体上の5bp欠失とキメラ子8のGFP欠失)さて、問題はこの時のキメラ子9匹のDNAが残されていることです。桂報告書10Pです。
>>
(調査結果) 1)Article Fig.4 と Extended Data Fig.7 に 129/Sv×B6(CAG-GFP) F1 マウスから作ら れた STAP 細胞由来の 2N キメラができたこと、さらに germline transmission により、 このキメラの子ができたことが報告されている。 小保方研のフリーザーに「カルスキメラ子 1」~「カルスキメラ子 9」と書かれた 9 本の DNA 試料があり、2011~2012 年の CDB 若山研では STAP 細胞を「カルス」と呼んで いたことから、これらはこのキメラの子の DNA と考えられた。実際に、小保方氏への聞 き取り調査により、これらの試料は Article Extended Data Fig.7 に出てくるキメラの 子から小保方氏が抽出した DNA であることを確認した。若山氏の実験ノートでは、この キメラの作製は 2012 年 1 月終りから 2 月はじめにかけて行なわれていた。
9匹なんです。ジャームライントランスミッション実験時に親として選ばれたキメラがWild type(Wide typeはタイプミス)を除いて9匹でした。でもこれはキメラのF0です。キメラ子という以上F1でしょうね。つまり35匹生まれた中の9匹でしょう。この検証結果が桂報告書のスライドにある。こちらはBCA報告にはないものです。以下です。
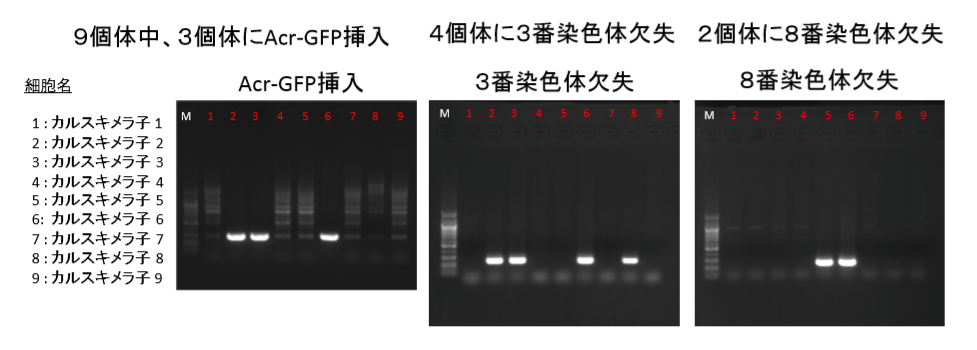
まず、3番染色体にAcr-CAGの入っていたカルスキメラ子は2,3,6です。このGFPが入っているのは岡部マウスです。岡部マウスというのはB6マウスです。F1マウスのB6側の3番染色体にこのAcr-CAGが入っている。同じく3番染色体には5bpの欠失があってそれはカルスキメラ子2、3、6、8にあるのですが、実はこれもB6側の3番染色体なんです。同じく桂報告書のスライドです。
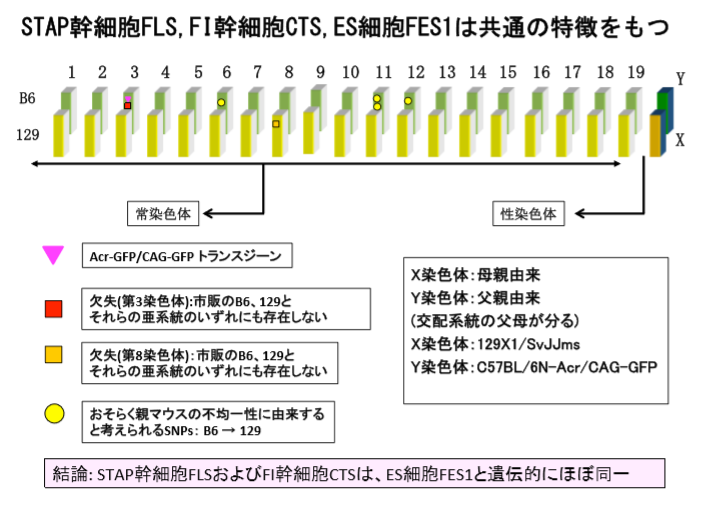
ピンクの下向き三角と赤の四角です。
B6側のGFPと欠失は同じ3番染色体上にあるので一緒に動きます。カルスキメラ子の8にはGFPがついてきていません。不思議です。
原因として通常考えられるのは減数分裂時の乗り換えでしょうか。2Nキメラのジャームライントランスミッション時に、できた子供の一つに、GFPが減数分裂時に相同組み換えを起こして別の染色体上に交叉融合して飛んでしまっていたものが居たということになる。しかしGFPはこの場合PCRで探しています。岡部マウスと分かってしまっているのですから既知のプライマーで挟んでます。別の染色体に飛んでいても存在していたらPCRにかかってくるはずですね。乗り換えではなさそうです。
因みに5bp欠失の見つけ方としては、場所はシーケンシングで分かっていますから、その場所の前後近辺のプライマーを作ってPCRにかけて挟むと長いのと短いのが出るので短い方が4つバンドになっているところが提示されているわけです。
GFPに関してはマウスコロニーにGFPの無いB6が飛び込んだのか、減数分裂過程、もしくは培養時の体細胞分裂過程でGFPの欠失が生じた等の可能性が考えられるが、5bp欠失は4つに共通していて1つだけにGFPが無いという現象がどの可能性を許すかという問題になる。GFPの有るマウスと無いマウスのどちらにも5bpの同じ欠失が生じるということは無いので、やはりGFPのあった細胞に5bp欠失が生じ、それをいくつか株分けして継代培養しているうちにその一つにGFPの欠失が生じたと考えるしかない。できたばかりのSTAP細胞を移植してこういうことは起きない。我々のntES仮説を支持している数々の証拠の中の一つの現象ではないか。
因みに太田ESは作っただけで使いもせずにすぐ凍結して、解凍するまでは時間は止まっていたので、それに培養変異があるなんてことはない。桂報告の小保方さんによる太田ES使用捏造結論でも、変異が入るなら小保方さんが解凍した後の話ですね。若山さんのntES化であっても、小保方さんの太田ES使用であっても、変異期間は同じ長さです。桂報告書の論理はとても非合理なもので科学者の書いた文章とは思えないところです。小保方さんは光っているマウスだけを9つ選んだと推定したが、PCRの結果は光っているのは3つしか選んでない。キメラ子自体は105匹生まれていてその中の毛色でジャームライン有になったもの35匹の中から9つ選んでいると考えたのですが、このPCR結果は105匹の中からランダムに9匹選んだことになる。それで35/105はほぼ3/9で計算上はあっています。
これらカルスキメラ子105匹のマウス背景は何であり得るかというと、元の2Nキメラの生殖細胞はドナーである129B6であるか、リシピエントであるICRであるかのどちらかです。毛色でキメラになっているもの同士の兄妹交配ですが、GFP蛍光する生殖細胞以外はどうであるかは実際には関係ありません。
配偶子は129とB6そしてICRとICRに減数分裂する。たまたまICR同士がメイティングしていたら子供はすべてICR/ICRが生まれる。129B6同士だったら129B6が2、129/129が1、B6/B6が1の比率で生まれます。ICRと129B6がメイティングしていたらICR/129が2とICR/B6が2生まれます。B6の入っているものだけが光る。
因みにキメラの子はキメラではありません。普通のマウスです。
岡部マウスは太田さんが岡部さんの元から若山研に来た2003年からあるはずのものです。彼は精子での研究に来ている。無論自分の研究のために若山研に持ち込んだものなので、それ以来若山研で維持されている。
「僕のマウス」の親の129ではとんでもないマウスコンタミがありました。岡部マウスに関してもコンタミ懸念は無いのか。マウス会社から別のB6を買ったことがあるのは記者会見で証言もされている。
ただ、手記によると若山さんは光る精子を集めていたと書かれているので、岡部マウスの交配は顕微授精で行われていた可能性もある。卵はCAGで光るし、精子はCAGで光らない代わりにアクロシンで光る。このやり方で継代し続けている限り、比較的マウスのコンタミは排除される。特に他の遺伝子コンタミは防ぎえないかもしれないが、GFPに関してはこれの無いB6は取り除かれてしまうのでGFPのないマウスは無いことになる。
桂チームは2010年に若山研で凍結された岡部マウスの受精卵を調べて3番の欠失は無いと言っています。どうして最新維持されている岡部マウスは調べなかったのか。小保方さんの研究のための実験は2011年から2012年にかけて行われた。2010年の凍結卵だけ調べては分かりません。こんなついでに調べられるような今の岡部マウスを調べないというのは結論ありきの線から外れそうなものはもともと調べないという方針だと疑われても仕方無いでしょう。
桂報告はFLSは太田ESのFES1だということにしてしまったのですが、FES1は2005年の論文で使われた129と岡部マウスを掛け合わせた129B6F1のntES-G1G2、もしくはその元を作った頃に、何の目的もなくただ同じマウスから受精卵ESを作ったと、本人によって物語られているもので、歴史的な証拠は何もないものです。本人は日経サイエンスの取材で細胞リストに書かれていると証言しているようですが、その開示はありません。因みに理研に提出されているntES-G1G2のラベルには129B6F1G1と129B6F1G2と書かれているのですが、理研の調査結果ではどちらもB6/129で親の雌雄が逆になっていました。太田論文には129/B6となっています。つまり論文記載背景とラベルは一致しているが中身が違っているわけです。
G1,G2と名前を付けたのが2008年かどうかは調べられていないから分かりませんが合理的推測では2008年です。先に作られたものに最初から2番と書かれているわけはない。中身が論文ともラベルとも違っていると分かっていながら証拠の信憑性確認をスルーするというのは詐欺犯罪に近い行為ではないでしょうか。
(培養変異)繰り返しますが、太田ESが2005年の論文のための実験中に作られていて2014年に調べられるまで10年近いから培養変異が入りやすいという理屈は無論嘘です。凍結されている間細胞の時間は止まっています。変異は培養回数によるんです。何日間培養され何回植え継がれたのかということはどのサンプルも調べられていません。そもそも実験ノートの提出公開は一部といえども小保方さん以外は行っていません。小保方さんの提出した実験ノートを公務員法違反行為と知りながらNHKに全コピー流出させた松崎GDを起訴もしないという、これも同時にそれ自体も公務員法違反である行為を犯していながら、小保方さんの実験ノートが提出されなかったから分からないなどと、とぼけたことを書いている桂報告を放置している理研は、追々追及を受けることになるでしょう。
FES1,2は2005/12/7に凍結されている。何の目的もなく作ったと太田さんが証言している以上、できたのち長く継代培養することはない。作成したのは12月中でしょう。ntESG2の凍結は2005/1/20です。論文を提出して既に2005/4/26にはアクセプトされている。その後8か月も経過してから何の目的もなくFES1,2を作ったと言ってる。余った129/Sv-terを使って実験したのかと思いきや、ここではX1を使った。若山さんが別途購入していたものを流用したのでしょうかね。ただ、その場合は2005年の論文とは何の関係もありませんね。何のためにntESG1,G2を提出したのか。報告書4P。
>>
また、STAP 幹細胞 FLS が、Acrosin プロモーター下に GFP を発現する Acr-GFP と、CAG プロモーター下に GFP を発現する CAG-GFP の共挿入を含むことが判明した後に、 過去に CDB ゲノム・リプログラミング研究チーム(以下「CDB 若山研」という)で作製さ れた Acr-GFP/CAG-GFP 共挿入 ES 細胞を取り寄せて解析したのが、下段の 4 種類の細胞で ある。アクロシンが出たから過去に岡部マウスを使った論文関係の細胞を出せと言われて太田氏と若山さんが理研に出したのが、FES1,2とntESG1,G2という書きざまですが、Ooboe さん提供データで明らかになっているように、これは桂調査チームより先に若山さんから東北大に解析依頼提出されている。むちゃくちゃですね。ガヴァナンスも何もあったものじゃない。
FES1は無かったはずのものです。報告書14P。
>>
ES 細胞混入のもう 1 つの謎は、ES 細胞 FES1 がどのようにして STAP 細胞研究時の CDB 若山研に存在したかである。ES 細胞 FES1 は 2005 年に当時の CDB 若山研メンバー によって樹立されたが、その後、研究に使わず、2010 年 3 月(CDB 若山研で STAP 研究 が始まる前)に転出した時に ES 細胞 FES1 の凍結保存試料を全部持ち出して CDB 若山 研には残さなかったとされている。当時の CDB 若山研メンバーへの質問状と聞き取り 調査、および関係者の実験ノートの調査でも、当該メンバー以外に ES 細胞 FES1 を使 用した者は見つからなかった。 だったらFES1は使われていないと分かりそうなものですがね。
FES1には3番と8番に欠失がありましたが、8番は129側ですので、今はB6の3番の欠失です。FES1の3番の欠失はいつできたか。2010年の岡部マウスの凍結受精卵にはなかった。無いのにどうしてそれ以前の2005/12/7に作られている太田ESにB6の欠失があるのか。それは培養時にできたのだという解釈ですね。培養は2004年の実験中と、2007年の解凍後の再使用時の培養の二度です。何回植え継がれたか調べられていない。一方、FLSやCTSは2012年の初頭から秋口までと笹井研での再解凍培養の二度です。こちらより太田ESの培養期間が長いとも思えませんね。どちらにも3番の欠失は起こり得るんです。そしてFES1と称してFLSを中身に入れ替えたらこんなことは当たり前の結果です。SNPs近似はむしろそれを指摘しています。これは和モガさんがずいぶん以前から指摘しているし、佐藤さんの本でも書かれている。
この3番の欠失は当然ですがテラトーマからも出ている。
>>
(f) STAP 細胞から作製されたテラトーマは、ES 細胞 FES1 に由来する可能性が高い
(調査結果) Article Fig.2eと Extended Data Fig.4a-c に登場する STAP 細胞由来のテラトーマは、 いずれも Oct4-GFP+細胞の 7 日目細胞塊から由来したとされている。しかし、以下1) ~3)の検証結果に示す通り、このテラトーマは、 (1)Acr-GFP 遺伝子を含むが Oct4-GFP 遺伝子は含まないこと (2)ES 細胞 FES1 に特異的な 2 個の欠失が定量 PCR の解析で検出されたこと (3)組織切片の FISH 並びに染色体ペインティングで大部分の細胞に X 染色体と Y 染 色体各 1 本が検出されたこと(ES 細胞 FES1 が XY(オス)であるという事実と合致) が判明した。よって、これらの図に登場する STAP 細胞由来のテラトーマは、ES 細胞 FES1 に由来する可能性が高い。太田ESを渡されて騙されたと言ってる若山さんの嘘は既に証明しています。ESを渡されたら大きさの違いで誰にでもわかる。太田ESでないのにアクロシが出るのは最初から小保方さんに渡しているマウスが129/Sv-X1と岡部マウスとのF1だからです。
そして若山さんの当時維持していた岡部マウスには3番の欠失は2010年まで無かった。
無論、2010年の凍結卵にないものは2008年以前には無かったはずというのは、マウスがちゃんと均一に近交系に維持されている場合の話です。不均一ですとたまたまGFPの欠失しているマウスが選ばれなかったというだけの可能性もある。どうも「僕のマウス」の片親である129CAGホモのB6特異的SNPs領域が広くてマウスのコンタミが疑われている。白毛のマウスの中に黒毛が飛び込んでるとしたら、黒毛の岡部マウスに黒毛のGFP無しマウスが飛び込んでわからなくなったいうことはあり得ないことではない。
こういうコロニーの汚染はランダムに数匹抜き取って調べないと少ない数では分かりませんが、そもそも岡部マウスのコロニー自体は調査すらされてない。自家繁殖なんてさせてる事自体がどうなのか。もっとも、若山ラボの研究目的にとって大きな障害ではなかったということはあり得ますね。そんな研究室に、マウスのコンタミなんてあってはいけない小保方さんの研究が参加してきてしまったという予期しない事情があったということも推定される。でも所詮マウス管理、なってないんですよね。
マウス自体にGFP欠失の有るものが混じったとしたらそれは、2010年以前と以後と両方あり得る。
しかし、まあ、時々顕微授精もしていたとして、結構長く自家繁殖もさせているから、GFPがヘテロに入っていたマウスがあったという蓋然性もそれほど高くはない。すると培養時にGFP欠失が起きたと考えることもできる。体細胞分裂時に起きた欠失ですね。
この場合は酸浴中、酸浴維持培養中の7日間、そして、この細胞の核を移植してntES化して後の培養中、更にその維持培養中に生じたと考えることになる。FLS樹立と同時に行われた2Nキメラでしょうね。テラトーマに最初のF1の幹細胞を注射してしまっていますから一からやり直したとすると日程的には以下のようになる。
2012/2/2***若山さんが129/B6-Act-CAGの赤ちゃんマウスを小保方さんに4,5匹渡す。
2012/2/8***小保方さんがSTAP塊を若山さんに渡す。
2012/2/9***若山さんがリシピエントの除核卵に小保方細胞核を入れたクローン胚をたくさん作る。
2012/2/13***数日後クローン胚が胚盤胞期になったところでインナーセルマスを取り出してntES培養を始める。成功率は1割程度。増殖成功しているシャーレはそれぞれ雌雄どちらかである。
2012/2/20***培養一週間後で増殖確認できたものをキメラ胚に入れる。いくつかのシャーレから数個ずつを混ぜて一つのキメラ胚に20個程度入れる。
2012/2/25***いくつかのキメラ胚のインナーセルマスを取り出してES培養し、幹細胞化する。これがFLSのライン。小保方さんに8ライン渡して増殖率実験を行わせた。120日後、2012/6/25前後に実験を終えて凍結されたものがP40のFLS1-8のサンプルである。
2012/3/10***20日後に2Nキメラの出産がある。若山氏実験ノートに2012/3/11とある。
2Nのカルスキメラ子8にだけGFP欠失がある。
ではこのGFPの欠失は何時起きたか。すでにテラトーマにはAcr-CAG-GFPが見出されている。しかし、テラトーマの場合はたくさん注入するので、作られた1ラインに欠失があっても他のにGFPがあれば検出される。問題は翌年の実験であるカルスキメラ子の8番だけにGFPの欠失が生じていることです。
我々はntES化されていると見ている。酸浴細胞をクローン胚に入れてntESしたいくつかのシャーレの中からランダムに取り出してキメラ胚に入れているとみている。そのシャーレのどれかでGFPが欠失したものができたと考えることはできる。その際はもう一つの欠失は岡部マウスに先にできていたと考えるしかない。2010年には無かったがその後できたと。顕微授精を常時行っていてもありうる。この場合今の岡部マウスを調べないと分からない。なぜ調べなかったか。若山さんが捨ててしまっていて2010年のものしかなかったか。
(不動の定点)これを考えるときに不動の定点があるのと無いのでは天と地ほどの違いがある。我々は今小保方さんによる既存ESのコンタミなんて無いのだと分かっている。ここが押さえられるまでは本当に悩まされたが今や、STAP cells are derived from ES cellsなんてトンデモだと分かっている。この表題だけならntESであることも含みうるが内容は太田ESと学生のGOF-ESと若山さんの「僕のマウス」ESを使った小保方さんによる捏造という結論になっている。大間違いですね。
もう一度貼り付けましょう。ニーチェはこういう時よく「驢馬どものための注」と書くのを好んでました。
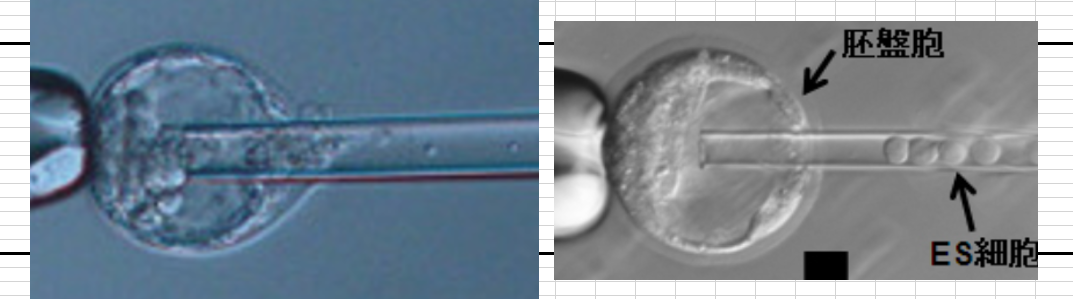
左は最初の実験時に小保方さんの細胞をナイフでカットして胚盤胞に移植している写真です。若山さんが行ってる写真ですから、細胞塊は小保方さんが渡しているものです。左は通常の受精卵ES細胞です。大きさが違う。
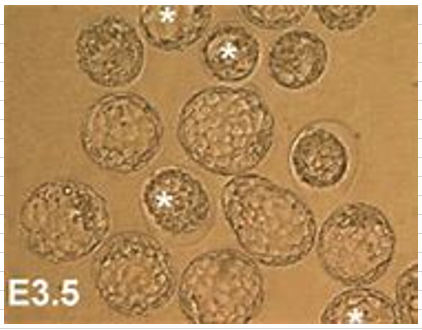
少々胚盤胞の大きさが違っても影響しない。以下はマウスの胚盤胞期である3.5日目の解剖取り出しされた胚盤胞です。この後ハッチングと言って外側のたんぱく質の膜を脱ぎ捨ててエピブラストになってから子宮に着床しますから、前段階で外膜が柔らかくなって全体が1.5倍くらいに急激に拡大します。そして直後にハッチングする。このハッチング前の期間のExpanded Blastcystと呼ばれる胚をキメラ胚として使います。大きさが違うのは3.5日目で検討つけて取り出しますから、1匹に20個ほどある卵の段階が微妙に違うからです。一番小さいのは胚盤胞になり立てで、一番大きいのがハッチング直前です。この中から適当な大きさの径のパイプで胚盤胞を集める。こういうのはキメラ作成のプロトコルに説明されていますね。
実験には100個とか200個が使われますから一番数の多いサイズで集めて、集める過程で人の目で選択しているからやや小さいのも入りますね。入れる胚盤胞が少々小さくてもESの大きさと小保方さんの細胞の大きさの違いが分からないなんてことはありません。
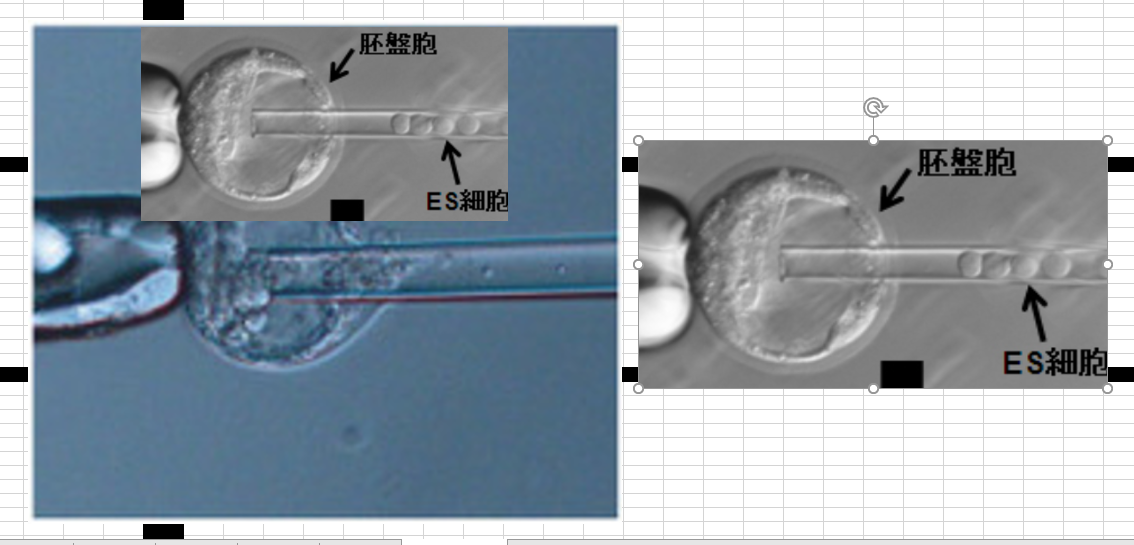
3.5日の胚盤胞写真で一番大きいのと一番小さいのは直径で倍違いますね。一番大きいのと一番小さいのがたまたま選ばれている確率はとても低いですね。みな同じ作業していますから平均的に一番多い大きさを集めている。でも両極端比較もしておきましょう。
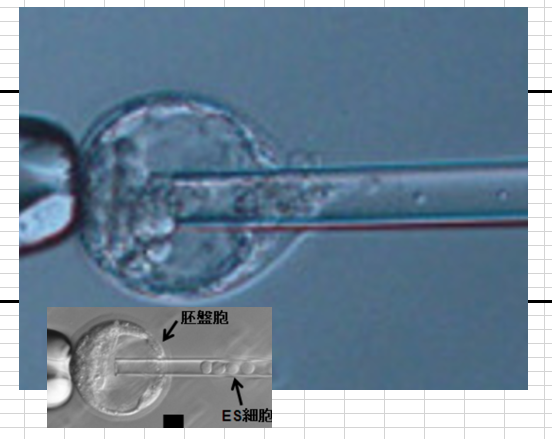
まだES細胞の方が大きいんですね。何年ESを見続けているのだということですね。小保方さんの細胞は基本リンパ球由来だから小さいんです。
直径で1/2、面積で1/4、体積で1/8です。同じ大きさにしようと思ったらESの写真はとてつもなく小さな胚盤胞を選んでいたということになりますね。以下のくらいかな。

こんな小さな胚盤胞を選ぶ人いないでしょ。そもそもSTAP細胞とES細胞の大きさは小保方さんが論文に写真を貼付している。彼女は形の比較を見せるために写真を同じサイズに調整していますから気づきにくいですが、左下にスケールがついているんですね。それを合わせたのが以下です。
ナイフ切り分け写真は後からのデモンストレーションとして撮影し直したかもしれませんが、彼は記者会見で質問に対してナイフ切り分けで入れた最初の実験でキメラができたんですと答えている。キメラの成功した時、彼はこんなに小さな細胞の幾分大きめの塊をナイフで切り出して、ES細胞の移植時より大きめのパイプを自分でバーナーで炙って引き延ばして、この細胞塊を移植したんです。そう自分で言ってるんです。もう一度貼り付けましょう。
失敗続きだったころ、彼はSTAP細胞の小さな粒に合わせたガラス管を作っていました。パイペットの作り方の図示は以下です。胚盤胞を左側で吸引して支持しているパイプをホウルディングバイペットといいます。カッティングニードルというのは若山さんが細胞塊を小さく刻んでいるあの針で、核移植の時にも使います。インジェクションパイペットが中にSTAP細胞やES細胞が入っているものです。


若山さんはSTAP細胞をトリプシンでばらして挿入している時はとても細いパイプを自分で作っていたことになりますね。彼のラボはntESキメラは常時作っています。受精卵ESもntESもその遺伝子解析的な性質は違いますが、大きさは同じです。普段ntES細胞は移植のためのパイプをいつも作っています。STAP細胞の時だけとても細く作る。細胞の大きさの違いを知らないということはありえませんね。
そして次に塊で入れたらその時に成功したから大きさについて意識していなかったといったんですがね。その後もその後も何回同じことしていることになるのでしょうかね。細胞の大きさ自体がかなり違いますよね。パイプとか胚盤胞との相対的な比較だけではありません。本当に何度もやっていたら大きさの違いに気づかないことはないでしょう。笹井さんがはっきり分かると言ってますね。
ナイフ切り分けなんてそもそも何度もはやってないと思いますね。最初の実験時だけですよ。
太田ESでないことが分かったら、ではなぜAcr-CAGがでるのか。それは若山さんが岡部マウスとのF1を使っていたからに決まってます。これはFLSを作った時、小保方さんに「僕のマウス」を渡したとという証言が虚偽だということを同時に証明しているんです。
手記208-209P。
>>
・・・若山先生は光る精子で実験をしていました・・・Article Extended Data Figure 7-bと7-cはSTAPキメラです。でも、若山さんの2011年11月までのキメラ実験失敗、検証再現実験で丹羽さんと小保方さんの作った酸浴細胞を清成さんがナイフ切り分けでキメラ胚移植実験した失敗結果からの我々の判断では、小保方酸浴細胞からキメラはできません。
従ってこのキメラはntESキメラだと考えるのが最も自然な推測です。
- 2019/09/05(木) 21:43:54|
- AC129
-
-
| コメント:0




























