学氏がGLSに関してブログに書かれている。私も今自分のブログでこの件に関して考察中ですが、事前にここに学説に関する批判を書いて置きます。
SNP調査などしない方が、ES説にとって都合が良いかもしれず、評価を後に残したのか?
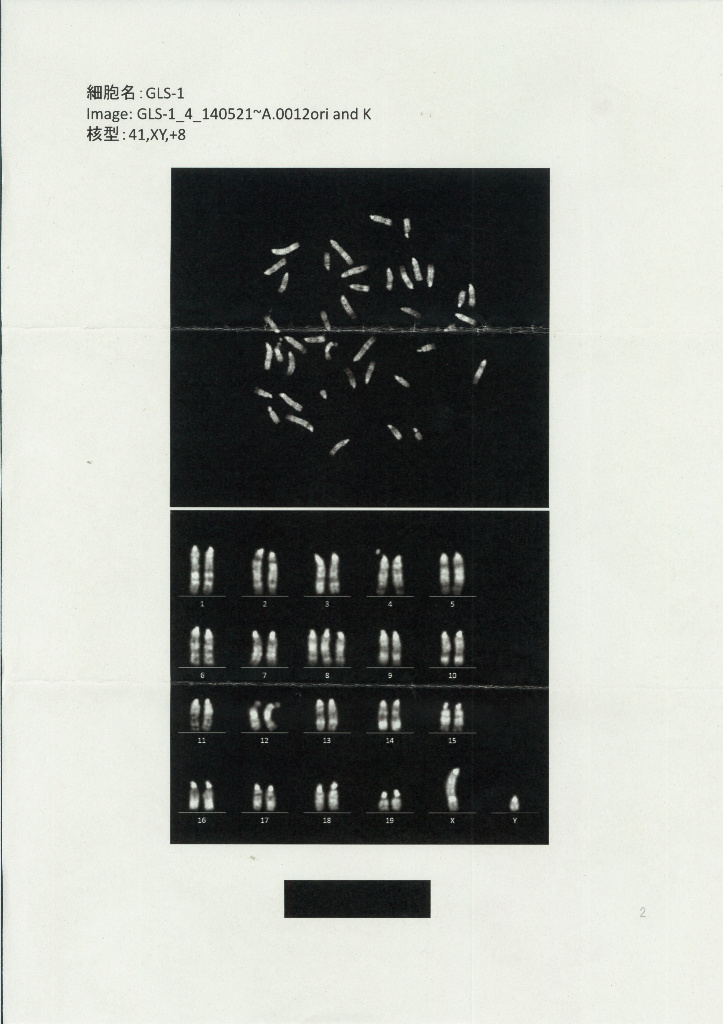
2019/09/17 万能細胞 iPS ES STAP GLS1とGOF ESは前者は小保方さんが若山研に自家飼育されていたGOFマウスを渡されてそのリンパ球を使ってSTAP細胞を作り若山さんに渡したものです。その後若山さんが何らかの手法で幹細胞化したと称しているもののうち理研側に残されたものの中から丹羽さんが性別確認のために2014/4/22に1と11を持ち出しています。木星リストの28と38番です。そのうちの1との比較です。
GOF ESというのは2010年の春から秋にかけて当時ラボに研修に来ていた糸井さんという学生さんが練習のために、やはり、若山研で飼育されていたGOFマウスを使ってntESをたくさん作っていたものを小保方さんが頼んでコントロールのESとしてシャーレ一皿を譲り受けていたもので、木星リストの102番のことです。2013/3/31までの若山研の引っ越し作業中に若山さんはMTA無しでこのサンプルの全てを小保方さんや第三者の立会無しに整理していますから、中身が当時のままであることは保証されていません。所謂、竹市さんが当時コメントした「細胞の出所が分からなくては特定の結論を導き出すことはできない」という忠告通りになってしまっているものです。警察の捜査であれば当たり前の事前の証拠能力の調査が全く行われないままに恣意的に調べられているサンプルです。お粗末としか言いようのないもので、こんなレヴェルで博士を称しているなんて、専門知識はともかくとして、そもそも基本的な常識は無いのかと一般人達に笑われているような調査です。
どういう時に、STAP細胞がES細胞から作られたと言えるかについて、桂報告書に書かれている。
特に、GLSの時に、条件が整っているので、一番、理解しやすい。
また、ES細胞へのすり替えとか、最初からES細胞が使われたとかでなく、桂報告書で、なぜ、“混入”との言葉が使われたのか?
ここに疑問を感じた人はいないのだろうか?STAP幹細胞はSTAP細胞から作られたと言われている。論文にそう書いてあります。嘘か本当かは分かりません。論文に書いてあるのですからその通りなのが普通は当たり前です。でも、残されているSTAP幹細胞と言われているものは、当時の理研若山研内にあった既存の受精卵ES(太田FESもしくは若山F1ES)、もしくはntES(糸井ntES)と同一であると証明されたがゆえに、STAP"幹細胞"がES細胞から作られたと桂報告とBCA報告は判断した。そして、STAP幹細胞は若山さんが小保方さんから受け取った細胞で作ったと証言しているからSTAP細胞は既存ES由来であったと世界にこっ恥ずかしい論理をさらしているわけです。無論裁判所でこんなこと言ったら裁判官からアホな事言うなと言われますよね。若山さんが小保方さんから渡された細胞をそのまま使ってSTAP幹細胞を作ったという主張は若山さんが自分で言ってるだけで何の証拠もない。恥ずかしすぎる論理です。小保方さんから渡された細胞をそのまま使ったのだという証拠を出さないと、STAP細胞が既存の受精卵ES(太田FESもしくは若山F1ES)、もしくはntES(糸井ntES)であったということはできないですよ。こんなことど素人が注意するようなことですかねえ。和モガさんも早くから指摘されていますよね。
竹市さんはテレビ会議で、「細胞の出所が分からなくては特定の結論を導き出すことはできない」と忠告して、野依さんも同意したというのに、こんな当たり前のことが実行されなかったというのは、理研にはノーベル賞受賞者とその候補者の二人しかまともな知性はいなかったということなのでしょうか。これはたぶん高校生レヴェルなら既に判断のつく問題ではないでしょうかね。無論嫌味を言ってるんですよ。私は。
小保方氏が故意にESを使おうとしたなら、どこでどうやって混ぜたの?について、さまざまな推論になる。最初から、ES細胞を使った方が確実などとの憶測も飛んだ。風が運んだ説、めったに起きない実験上のミスがたまたま重なった説とか、いろいろに言われた。そこで、桂報告書について、“混入”と表現された理由に注目して読んでみよう。そういういろんな説は聞いたことは有りません。小保方さんが既存ES細胞を渡したのだとマスコミは言い立てましたよ。桂報告とBCA報告は事故コンタミか意図的コンタミかは分からないが残されている幹細胞は既存の受精卵ES(太田FESもしくは若山F1ES)、もしくはntES(糸井ntES)であったと判断しているんです。そして常識的には事故の蓋然性は低いがゼロではないと言ってるんです。
この
事故コンタミの可能性に関しては太田ESが使われたと主張されている限りにおいては絶対にない。蓋然性ゼロだと私が論証しています。太田ESは理研若山研には無かったはずのESです。仮に置忘れがあったとしてもそれを解凍した人は故意です。
凍結細胞というのは解凍しないと使えないんです。こんなことにも気づかない桂チームは高校生以下だと分かりますよね。若山研内で誰一人太田ESのことは知らないと答えているんですよ。自分で太田ESが使われていると主張してるんですから、無かったはずで誰も知らないと言ってる太田ESを解凍して人がいる以上、嘘をついている人があることになるのは明確ですね。意図的に解凍した人がいないと使われようが無いが、誰も知らないといってるんだから意図的に解凍した人はいないのだと主張しているのが桂報告です。日本の恥です。全国の若人よ。こんな組織に入るんじゃないぞ。世の中にはまともな企業がたくさんある。
“混入”は、悪意のある故意以外でも、実験のたびに実験者も気づかないままで起きる可能性がある。桂報告書のこの部分を読むと、そうしたことを念頭に文章が書かれていることがわかる。
大事なのは、桂報告書は、実験ミスの可能性を念頭に入れている点ではないだろうか?違いますね。この事件で事故コンタミはない。このことは上記したこと以外にも
細胞がマウス背景に応じて使い分けられていることから古田氏ですら桂報告書に対して疑義を出しているくらいです。彼女の場合は小保方さんがやったのだと間接的に指弾する意図なんですね。でもそれは後で考えることにして、ここでは誰が犯人であるにせよ、既存ESコンタミとする以上は事故コンタミは無いのだと知らねばなりませんね。もし、既存ESのコンタミでは無かったと主張を訂正するのであれば、その時には初めて事故コンタミがあったのではないかと問い直すことになる。
その時には桂報告とBCA報告は小保方さんと全世界にまず謝罪しなければなりませんよね。既存ESのコンタミであるとしたままで、事故コンタミがあったかもしれないと強弁することは許されない。まず謝れ。どれだけ小保方さんがお前たちの間抜けな論理に苦しめられたか。謝りたくなかったらそのまま強弁しつづけていろよ。
まずは、GLSで、同一細胞性が十分に説明されている部分を確認しよう。
STAP 幹細胞 GLS の中から選んだ GLS1 と ES 細胞 GOF-ES 細胞において(桂報告書七頁 青字)
1)全ゲノム上の SNPs 分布(C57BL/6 マウス背景)が同一
2)挿入遺伝子の種類、コピー数、挿入領域の配列が同一
3)由来するマウスの性別(メス)が同一
4)X 染色体上の構造異常(大きな欠失+末端重複逆位接続)が同一
このうち特に大事なのは、SNPの一致であろう。
すなわち、遺伝子構造異常の一致と、SNPsの一致で、GLS1 と ES 細胞 GOF-ES 細胞は同じ細胞とみなされ、STAP幹細胞はGOF-ES 細胞から作られたと結論している。1)は当然ですね。元の背景マウスが若山研で自家飼育されているGOFマウスですから小保方さんの貰った学生のntESの元のマウスと若山さんが小保方さんにした赤ちゃんマウスは同じSNPs分布を持つマウスで、若山さんがそのままGLSにしたか、それとも核移植してntESにしたかはともかくとして、又MTAもせずに一人で細胞を整理しているわけですから若山さんがGLSを小保方さんのGOF ESとラベルしているチュープに入れ替えたかは、全て可能性として留保しておかなければならないが、元のマウスは同じものだと証明されているんですね。これは論文通りのSTAP幹細胞なのか、学生のntESなのか、若山さんの作ったntESであるかの問題とは何の関係もありません。元のマウスは共通だという証明です。
2)もOct4-GFPのことで、自家繁殖させているGOFマウスだということです。1)と同じマウス背景の同一性証明に過ぎない。
3)はDORAさんブログで昔検討された問題で、丹羽さんが持ち出して核型解析させオスだとされた。常識的に一見すればオスですね。
GLS-1です。
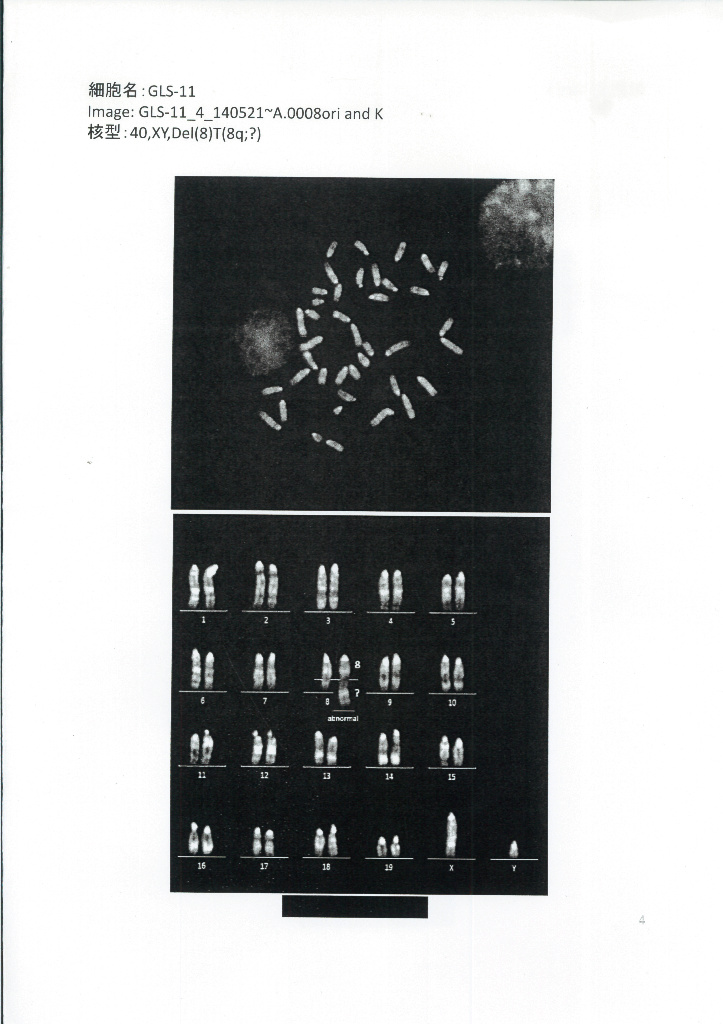
GLS-11です。
若山さんは6/16の記者会見で全部メスだと報告しましたね。

これは若山さんの説明で山梨大で13株の性別を調べたと証言されている。山梨大に保管されていた細胞株が全部メスだったということです。対して丹羽さんが調査に出したのは理研に残されていたものです。これは核型解析で1も11もXYと書かれていてオスの判定です。この後理研の調査結果と食い違っていたために若山さんは7/22に訂正した。
Y染色体に欠損があって用いたプライマーにかかってこなかったので無いと誤認してメスと判断したという。若山研では核型解析ではなくてY染色体の有無をPCRで確認したんですね。
そして、この結末はどうなったかというと桂報告でメスということになった。
10/21に松崎氏が持ち出して全株調べ直したんですね。その調査結果は以下でしたね。
>>
The STAP stem-cell line GLS1–13 was reported as established from STAP cells prepared from genomic Oct4 fragments (GOF) mice (B6 background) carrying the Oct4-gfp transgene[10] in 2012. All these cell lines have a large truncation with a terminal inverted repeat in one of two X chromosomes (Extended Data Fig. 2a). An identical X chromosome was found in GOF-ES, an ntES cell line established from GOF mice in 2011, but not in parental GOF mice. It is unlikely that such a peculiar X chromosome abnormality would occur independently, strongly suggesting that the GLS lines were derived from the GOF-ES.strongly suggestingというところに書き手の間抜けさが暴露されていますね。
サンプルは入れ替えられているんです。入れ替えられてないということを先に証明しないと裁判にすらならない。因みに全解析されているのはGLS-1だけですね。他はPCRなのか、シーケンシングなのか。注は*The line subjected to WGS is indicated in parentheses in cases in which several sublines were established for one cell type. Other sublines were confirmed by PCR and sequencing.となっている。
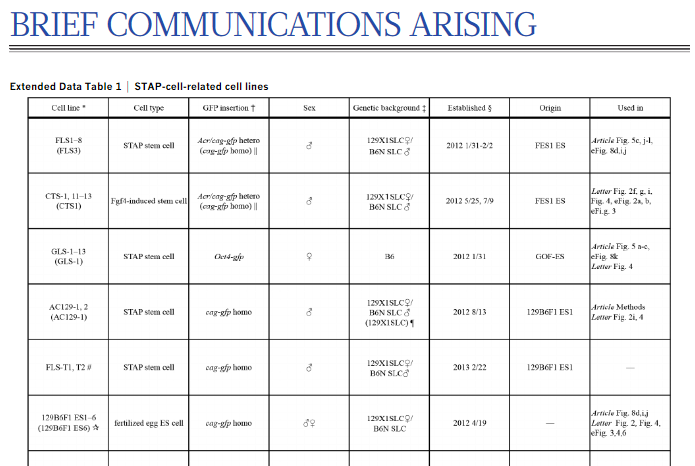
ともあれ、メスであるかオスであるかはどうでもいいんですね。もともとオスメス混ぜられていたSTAP細胞からどうして片方の性の細胞だけが出来てくるのかという疑義です。
既存ESコンタミでなければ若山さんのntESしかありませんね。どちらかはわかっていません。
そもそも若山さんのntESの可能性なんて調べられていない。間抜けも極まっているんですね。そもそも科学の専門家だというだけで捜査にはど素人以下の人材しかいないんですから呆れかえってしまいますね。一般人だって捜査の常識程度は知ってるぞ。テレビで探偵もの見てるしな。はは。野依さんと竹市さんが意見してるんでしょ。会議を主導した馬鹿は誰だ。日本ってここまででダメになってるのか。
ま、そういうことで、性別の件から4番目の検査結果が突然出てきた。今度はY染色体ではなくてX染色体に欠失と重複があるという。これは後に全部調べて13株すべてにあった。
若山さんの言ったY染色体の欠失はどうなったのだ。Y染色体に欠失は無かったのなら当初のメスであるという発表は正しかったということになる。というより、
Y染色体は無かったのだ。無かったのにどうして欠失があったと言ったのか。調べもせずに適当に話を合わせているだけだと知れる。若山さんが山梨大で保持していたGLS1~13はメスだったのに、理研側に残されていて、丹羽さんの調べたGLS1と11、そして後述しますが、小保方さんが論文に掲載しているGLSのどれか一つの計3つはオスだったということになる。ところが、松崎氏が後に理研側の全株を調べ直したらメスだったと言ってることになる。とんでもない矛盾です。計3つが一見オスにしか見えないのになぜすべてメスだということになったのか。
短いY染色体と見えるものはX染色体の大きく欠失したものだと解釈しないといけなくなるが、この染色体の欠失に関しては何の説明もない。以下の説明にある末端逆位重複接続は長さの問題とは無関係なものです。(大きな欠失)の証明がない。
「最も強い疑問は、第五章ですでに述べたように、GLSの性別が逆転(オス→メス)していることである。調査委がこの性別の逆転について何の説明もせず、また性別の逆転の理由なると思われる「大きな欠失」について具体的に触れないのも不自然である。」(佐藤貴彦著 『STAP細胞 残された謎』 155P)4)X 染色体上の構造異常(大きな欠失+末端重複逆位接続)が同一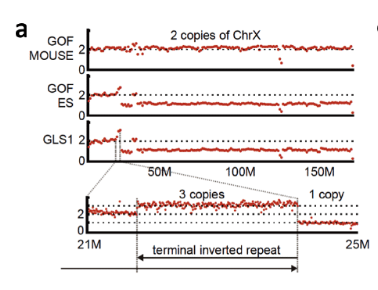
ともあれ、混入根拠に関しては、これが決定的なんですね。同一性の証明としてはこれだけです。他は同じマウスを使ってるということだけですから同一性の必要条件に過ぎなくて十分条件ではない。この4)こそが十分条件なんですね。少なくとも調べたGOF ESとGLS1~13は同じものだ。つまり
①GOF ESを小保方さんが若山さんに渡したからGLSが作られた。
②GOF ES ラベル容器を若山さんが洗い出してGLSを入れた。
①でないことは既にES細胞の大きさの違いから笹井さんの指摘が証明されている。②なんです。小保方氏が若山氏からわたされたマウスの状態では、X染色体の異常はないと思われる。引用するときは全部して欲しいですね。
>>
5)マウス個体で X 染色体上に上記のように大きな構造異常が生じた場合、その染色体 は世代を超えて安定に維持されないこと
6)ES 細胞 GOF-ES の元となった親の GOF マウスには、X 染色体構造異常が認められなか ったこと
7)GOF マウスの SNPs 分布が、STAP 幹細胞 GLS1 および ES 細胞 GOF-ES の SNPs 分布と異 なっていたこと
8)STAP 幹細胞 GLS1 以外の全ての独立な GLS 株でも、STAP 幹細胞 GLS1 と同じ X 染色体 上の構造異常が見つかったこと が判明した。 マウスにX染色体の異常が無いことは報告書に書かれています。思われるなんて、推測と報告事実を一緒くたにしないでいただきたい。
推測だが、培養途上でSTAP凝集塊が増大していく過程で、質の違う細胞が一緒に凝集塊に加わるとは思えない。
となると、特殊な生存状況で凝集している細胞に直前に“混入” するとなる。
この場合は、Ooboe氏らが指摘するように、その形態から若山氏が気付かないわけがない。
注入する時は、凝集した細胞を個々に若山氏は見るであろうと思われるからである。思いだけでは証拠になりませんね。
次の桂報告書8頁の中に、①②の数字を学とみ子が入れたが、ここは、報告書が、ES細胞が混じってしまう可能性のある時点を指摘している。
混入の危機は、この二時点である。
①は小保方氏の作業で、②は若山氏の作業である。
本調査委員会では、「STAP 幹細胞 GLS と ES 細胞 GOF-ES は同一由来の細胞 である」と認定した。
また「GOF マウスから ES 細胞 GOF-ES が樹立された過程で X 染色 体上の構造異常が生じ、①GOF マウスから STAP 細胞を経て② STAP 幹細胞 GLS が作製された 過程でこの ES 細胞 GOF-ES の混入が生じ、それを用いた実験結果が Article の Fig.5 お よび Extended Data Fig.8 に示された」と結論づけた。以下のArticle Figure 5-aのDAY7の画像は小保方さんは作れないのだから写真を撮ったのは若山さんです。bはSTAP細胞は塊で培養維持されていて増殖しないが、STAP幹細胞はバラバラにしたものから増殖しているという増殖能を獲得したという証明画像。cは増殖率表ですがES細胞の実験はどこかからのデータを使用していて行われていないが、STAP細胞とSTAP幹細胞(FLS)の増殖実験は行われていて40パッセージのFLS1~8が残されている。我々の仮説ですとntESなのでこの結果は当たり前です。
Extended Data Figure 8は以下です。
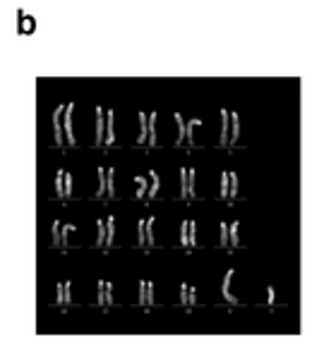
Extended Data Figure 8-bにはGLSの核型解析結果が添付されているが、丹羽さんが検証したのと同じ
オスですね。トリソミーも無い。
dはメチル化実験結果でキメラが出来ているので脱メチル化していなければならないのに、してないから、データを何度も取り直して実験結果を捏造したとされているものですが、そもそもキメラが出来ていると騙されているのですから、こんな状況に置かれたら誰だってもちゃもちゃやることになるでしょうね。いや自分ならやらないという人は、若山研に世話になっておきながら、しかも、できたと言ってくれている人のことを捏造者だと糾弾しなければならなくなるでしょうね。そんなことってできる人いるか。しかもポスドクの客員の身分で。ただ、一人の科学者としては変だと気づかないといけないのではないかな。気づいてもっと早く去るべきでしょうね。気づいてないから理研の呼び戻しにも応じましたね。
①で混入が起きてしまう理由は、まれには器具の汚染があるだろうが、可能性は意識的な“混入”である。
(ただし、学とみ子は、誰にしろ故意のES混入説には否定的)
②の場合は、意識的にES細胞を混ぜるのではなく、実験手技を通してのミスである。
故意でない混入の方が、可能性が高いと思う。
いづれにしろ、桂報告書(青字)はこの二点において、“混入”を疑った。
だからこその“混入”との言葉なのである。桂報告書はまず(GOFマウスからES細胞GOF-ESが樹立された過程でX染色体上の構造異常が生じ)たと言ってる。学生の糸井さんが作製した時にできたと想定していますが、まず糸井さんはたくさんシャーレを作っていて小保方さんはその中の一つを貰ってGOF ESと名付けて継代し、凍結ラベルしていたということです。糸井さんが当時作った細胞は糸井さんがどこかに保管しているのかどうかは報告されていない。調べたかどうかすら分からない。糸井さんが作った時に入ったなんてことは全部を調べないと分からないことで、どんな株分けをしたのかもわかりませんね。たまたま小保方さんの貰ったものに遺伝子異常があったのかどうか。ntESというのはたくさんのクローン胚から成功しているものだけをES化して更に成功したものだけが樹立とされるものですから、由来が皆違うものです。マウスにはこの変異は有りませんから、核移植から先の培養変異です。
学生のntESというのはリンパ球由来ではないはずです。恐らく尻尾の組織細胞か何かでしょう。それに対して小保方さんが渡したはずのSTAP細胞はリンパ球です。GLSのLはLimphocyteです。私の仮説でも使われた細胞はリンパ球の酸浴細胞核使用ntESなんです。GOF ESでもGLSでもいいが、これにTCR再構成があるか無いかを調べると、あれば少なくとも学生のntESは使われていないと証明されるんですが、この調査はなされていない。公共データ登録されているものもGLSの解析はされていませんね。
糸井さんに使った組織は何であったか確認する。そしてGLSもしくはGLのTCR再構成確認をする。それだけで証明されることすら行ってない。決めつけ調査なんですね。小保方さんがESを混入したという決めつけです。これを言い出したのは若山さんです。自分の渡したマウスで無いと言い出した。
シャーレ全部が変異した細胞で満たされるということはありません。一部に変異集団があるということですから、どこまで分割継代して行っても正常な細胞は存在している。変異が入った細胞だけが増殖力を増して全シャーレを支配するバイアスがあるという保証もありません。一つのシャーレを調べたというのはその中の全部の細胞をすりつぶしてDNAを断片化して、シーケンサーで読み取るときに異常の有る断片が見つかったということです。調べられたのはGOF ESとGLS1~13で、全てに異常が共通していたと、桂チームは言う。
これまでひどいレトリックばかり弄しているからそもそもこいつらの人間性が信じられるのかと疑うと、実験結果すら疑わしいということになりますが、まあ、科学者として最低の矜持くらいはあるだろうと判断して実験事実は信じるという立場で検討しているわけです。
変異がいつ入ったかに関しては、
①若山さんがGLSとしてntES化する過程(GL作成時点からの可能性が高い)のどこかで入った。核移植後すぐに入ると全体が変異集団になる。維持培養中に入ると、株分けの仕方にもよるが、一部に入ることになる。GLSは全株に入っている。無論GOFESは後にGLSに中身が入れ替えられている。偶然に同じ異変が入ることはない。
②①のケースと同様に糸井さんがntESを作った時のどこかの過程で入った。それを小保方さんが若山さんを騙して渡した結果、できてきた幹細胞は全部同一変異があることになった。
②はないんですね。もしそうなら若山さんは出来ないと小保方さんに訴えられた時に簡単に教えることができたことになる。人が違うとできないなんてことはあり得ない。小保方さんの前で小保方さんの作った細胞で幹細胞化させることはできなかったということです。結論は①しかありません。学さんがおっしゃるような事故ESコンタミは無いんです。
[①GOFマウスからSTAP細胞を経て② STAP幹細胞GLS作製された過程]を前提にしてはいけませんね。それは片方の容疑者が言ってることに過ぎない。可能性は桂報告の推定だけではない。小保方さんの細胞核を使った別の実験であった可能性は考えもされていない。調査のやり直しが必要ですね。
毎回作られるSTAP細胞作成時、大きな遺伝子異常がおきれば、そこらからキメラはできてこない。
X 染色体上の構造異常やトリソミーなどを持つマウスは生存できないが、細胞実験には用いることができる。
ネーチャー論文にその細胞の写真が載っていてるのは、細胞としては生存が可能であるからと思われる。トリソミーは丹羽さんの調査に出したときの写真にありますね。GLS-1の8番染色体にある。でもGLS-11と論文のExtended Data Figure 8-bにはトリソミーはありません。
桂報告書8P。
>>
なお、STAP幹細胞 GLSには第 8 染色体のトリソミーがあったが、GOF マウスおよびES細胞 GOF-ES にはなかった。このトリソミーはマウスでは致死だが、ES 細胞でときどき 生じるもので、STAP幹細胞 GLS作製(GOF-ES混入)時または作製(混入)後に生じたと考えられた。 GOFマウスに無いのは当たり前ですね。トリソミーがあったら子供は生まれません。2014年の調査時点で、GOF ESにはトリソミーが無かった。しかしGLSにはあったというが、全ラインにあったかどうかはまたしても書かれていない。杜撰極まりない論理ですね。
でも丹羽さんの調べたGLS1には明確にトリソミーがある。11はトリソミーにはなっていないが同じ染色体に乗り換えが見られる。そして論文リヴァイズ時には何の異常も無いオスの核型解析結果がつけられている。理研の2014年時点での最終結論はメスです。
2014年時点でGOF ESにはトリソミーがなかった。GLS1にはトリソミーがある。つまり入れ替えられたGLSはGLS1以外だということまでは分かる。GLS-11には交叉乗り換えがあるが、GOF ESに乗り換えがあったか否かは報告されていない。2013年のリヴァイズ時点につけられたと推測される幹細胞化されたSTAP細胞の遺伝子異常が無いという証明写真は、GOF由来のSTAP細胞で無いとおかしいが、遺伝子異常は全くない。これがGLSであるなら、というよりGLSであるはずであるが、これには全く異常が無いので、異常の無いしかも、オスのGLSが小保方さん側に残されていたということになる。そして、オスであることは丹羽さんの検証とも一致している。
変なのは若山さんが山梨大で分析したGLS1~13なのであって、これはメスだと当初発表されたが、丹羽さんの検証結果と合わせるかの如くに、一旦Y染色体に欠失があってPCRにかからなかったと言う言い訳とともにオスとされたが、桂報告はそれを更にひっくり返してメスとした。『STAP細胞 残された謎』153P以降に問題提起されていますね。この問題は解決されていない。今、楠本さんがGLに関して問い合わせられていますから、そのうち何か情報があるかもしれませんね。
GLSでの評価がわかる人なら、どういう時に、STAP幹細胞がES細胞から作られたと判断できるかの基準が理解できる。
つまり、性の一致は当然のこととして、構造異常(重複、欠失、くりかえし)などの一致に加えて、一塩基変異も一致していることが大事なのである。遺伝子解析の専門家でない一般人でも理解できるように、懇切丁寧に桂報告書は書いている。ここを書いた人たちは、科学的判断をしっかり世に残したいと思ったのだろう。何かSNPsに関して誤解があるようですね。SNPsの一致とGFPの挿入位置の一致は若山研で飼育されていたGOFという背景マウスの一致証明で、そこから作られた、学生のntESか若山さんのntESか小保方さんのSTAPかの識別の証拠ではありません。遺伝子異常の一致こそが、GOF ES=GLSの動かぬ証拠です。つまり、
若山さんが中身を入れ替えているという動かぬ証拠です。
動物から細胞がつくられた直後、あるいはその逆の時も含めて、両者の遺伝子構成は一塩基変異も含め一致する。
一塩基変化も一致するので、細胞同時で、一方が他方から作られたことがわかるが、同時に、一塩基変異は、細胞がとられたマウスの遺伝子状況を反映している。一塩基変化も一致している。SNPsの基本の誤解ですね。近交系マウスですから20継代されて近交系マウスとして樹立された時点で、その時のマウスのSNPsは固定されます。近交系マウスはクローンと同じなので、樹立されたときに存在している何万か所かの特異的SNPsが全部ホモでかつ自分自身との交配ですから変化しないんです。だからGOFマウス由来だというのも、GOFマウス特異的なSNPsを調べてデータと一致していたら判明するわけです。近交系マウスですからその後交配を重ねても他のマウスのSNPsが混じり込むことはマウスコンタミでもない限りはありません。ただ、交配されるときに配偶子に新たなSNPsが生じることは有りますが、何万か所で同定されていますから少し加わったからと言って判別に難はありません。ある程度長く継代されて無視できないくらい増えた時には新たにネーミングを変えますね。
それから当然ですが培養変異は体細胞分裂にすぎませんから遺伝はしません。SNPsというのは代々積み重なってきているDNAの遺伝子変異ですよ。近交系マウスではそれが樹立時のまま固定されているというだけの話です。通常は両親から別々のSNPsを引き継いでいきますから刻々と組み合わせが変わる。近交系マウスでは自分自身と交配しているようなものですからそれが無いので、そのマウスの特異的SNPsがデータとして公表されているということです。
そして、その後に動物継代や細胞培養をくりかえし時間が経過してくると、相互に独自の塩基変異が起きる結果、その一致率に影響がでて来るのである。培養変異はSNPsとは直接関係ありません。
ここをふまえて、FES1の場合を考えてみよう。SNPs比較だけの話ですね。
GLSでの全一致の状況と比較すると、FLS3、FI 幹細胞 CTS1、FES1の場合は、そうした関係ではないことがわかる。同一性の決め手になるのは、偶発的に起きる一塩基変異の一致である。培養変異としての一塩基の変異を考えられているのなら、それは紛らわしい。SNPsというのは同一近交系マウスとして固定されている数万か所の一塩基性変異のことです。ここにジャームラインで加わってくるものとも違う。ただ単に培養中に起きている変異でまとまった遺伝子の繋がりではなくて一か所がDNA複製の時にコピーミスするというような話ですね。これは他の染色体単位での変異と同じに考えればいいので、基本のSNPsのどれからもランダムに乖離していくのでSNPsの一致率とは関係ありません。
FLS3、FI 幹細胞 CTS1、FES1、129/GFP ESは、共通の細胞に由来するとの考えは同じjだが、GLSの時のようにSNPまでぴったり一致しているわけではない。FLS3、CTS1、129/GFP ESの三細胞間ではSNPがほぼ一致だが、FES1とではSNPが少し違っていた。
桂報告書5頁(青)
FLS3、FI 幹細胞 CTS1、ES 細胞 FES1、および小保方 研で見つかった 129/GFP ES の、常染色体に存在する 129 ホモの SNPs が、突然変異、 あるいは遺伝的背景の不均一性によるものとしても、もしこれらの幹細胞がそれぞ れ独立に作製されたものであるなら、これらの 4 か所に共通の SNPs が観察される 可能性は低く、これら4種類の幹細胞が共通の細胞に由来することを強く示唆する。これは以下の近親率表の話ですね。
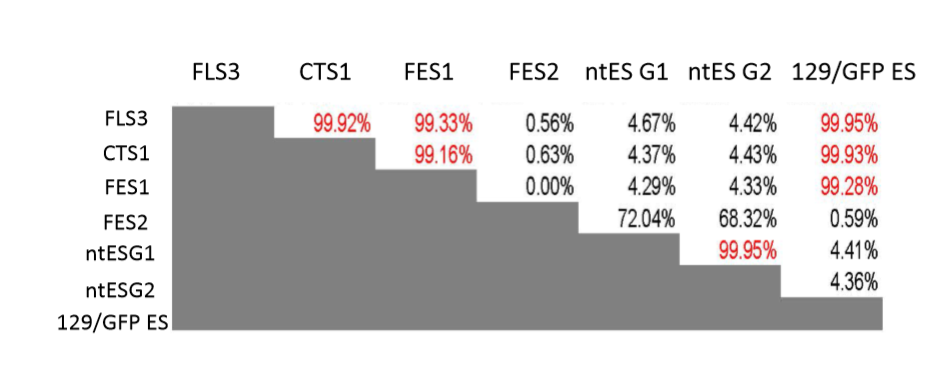
まず第一に混同しないようにしないといけないのはGOFマウスは近交系マウスですが、この近親率表に出ているマウスはF1です。近交系マウスではありません。129/SvとB6の交配種です。父親のB6はほぼ近交系マウスでしたが、母親の129/SvのCAGホモマウスは若山研でのマウスコンタミがあったらしくて、全く129/Svになっていないばかりか、それなりの近交系にもなっていなくてコロニーに不均一があります。完全な近交系同士でしたらSNPs分布は半々のグリーンになりますが、以下の通りなっていない。しかも不均一の証拠としてわずかな違いが出ている。
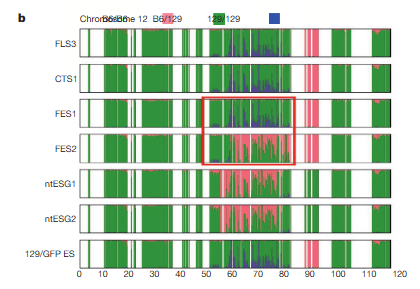
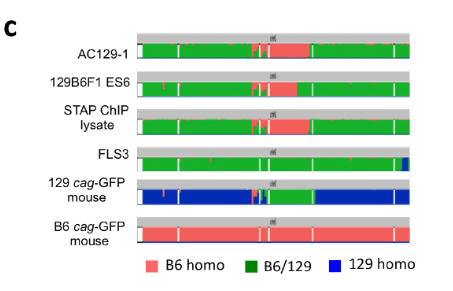 とくに、今回、FES1とFES2のSNPの異なる部分をあえてピックアップしてしまったために、ますます、FES1とFES2がそれぞれ、どの年度に作られた細胞に近いのか?がわかるようになった。
とくに、今回、FES1とFES2のSNPの異なる部分をあえてピックアップしてしまったために、ますます、FES1とFES2がそれぞれ、どの年度に作られた細胞に近いのか?がわかるようになった。
FES1とFES2において、他の細胞のSNPの一致率を比較することにより、FES1とFES2の作製時期は違うのではないか?の疑問である。FES1とFES2の違いは制作時期の違いではなく、129/Sv-X1と129/Sv-terの違いではないかと和モガさんが推測しています。ntES-G1G2はterです。FES2には似た分布箇所がある。これもサンプルの入れ替えが疑われているんです。
SNP調査などしない方が、ES説にとって都合が良いはずである。SNPs解析をしなければ既存ESコンタミ結論は導けなかったでしょう。ただその論拠に破綻が出ただけでしょう。
桂報告書がこれを書いたことは、STAP幹細胞がES細胞から作られたとの評価を、後の人に託したということではないのだろうか?小保方さんを陥れたのは桂報告とBCA報告です。何が後を託すですか。小保方さんをあんな目に合わせた奴らのために何を弁明してやろうとしているのですか。
- 2019/09/18(水) 08:49:14|
- 一枚報告補記
-
-
| コメント:0













