(2012年の実験日程)STAP樹立期間は7日、幹細胞樹立は4継代(12日)、メイティングは50日後、胎児は12日胎児、出産は20日で計算してみる。1,2日のずれはある。
まず、知られている情報通りに計算してみる。STAP幹細胞は論文通りSTAP細胞からのES培地誘導と仮定する。
1月23日 月 小保方さん帰国初出勤。
1月24日 火 129xB6-GFPの赤ちゃんマウスを雌雄取り混ぜて数匹渡された。STAP作成開始。
1月31日 火 若山さんFLS-1樹立培養開始。2N・4Nキメラ作成開始。
2月02日 木 若山さんFLS-2~8樹立培養開始。
2月09日 木 FLS-1三継代目。
2月11日 土 FLS-2~8三継代目。
2月12日 日 FLS-1四継代目。
2月13日 月 STAP由来2N・4Nキメラ12日胎児帝王切開取り出し。(Extended Data Figure7-d-下)2月14日 火 FLS-2~8四継代目。
2月15日 水 FLS-1~8の2N・4Nキメラ胚移植。
2月21日 火 STAP由来2N・4Nキメラ出産。(Figure 4-c,Extended Data Figure7-a-右、7-b)2月27日 月 FLS-1~8の2N・4Nキメラの12日胚の帝王切開。写真撮影。3月06日 火 FLS-1~8の2N・4Nキメラの出産。3月12日 月 西川氏本人主張TCR再構成アドバイス。(手記は4月のネイチャーリジェクト後)
3月22日 木 アニマルカルス命名。
4月06日 金 CDB若山研メンバーがFLSの4NキメラのDNA抽出。(出産から1か月後に当たる)
4月11日 水 STAP由来2N・4Nキメラメイティング。
4月19日 木 129B6FES1(僕のマウスES)樹立培養開始。(桂報告)(若山さん持ち出しリストでは5/25)(1~6まである)
4月24日 火 米国特許仮出願。
4月25日 水 FLS由来2N・4Nキメラメイティング。
4月27日 金 倫理委員会にて竹市所長の知るところとなる。
4月30日 月 4月中にネイチャーリジェクト。
5月01日 火 STAP由来2N・4Nキメラ子出産(2Nは兄妹交配 Extended Data Figure7-c 小保方氏が抽出した DNAあり、4Nは戻し交配?結果無し)5月15日 火 FLS由来2N・4Nキメラ子出産。(2Nは兄妹交配 Extended Data Figure 8-i 小保方氏が抽出した DNAあり、4Nは戻し交配 Figure 8-j)5月25日 金 CTS1 樹立培養開始。129B6F1ES(僕のマウスES)樹立培養開始。129B6F1TS(僕のマウスTS)樹立培養開始。(若山さんの持ち出しリスト)
6月06日 水 セル誌リジェクト。(TCR再構成の切り貼りレーンが使われている)
7月09日 月 CTS11~13樹立培養開始。
8月13日 月 AC129-1,2樹立培養開始。
8月21日 火 サイエンス誌リジェクト。
今度は我々のntES仮説でこの時に2011年の実験を最初から確認したと考えてみる。つまり、一からntESを作り直すとして計算してみる。矛盾はそのまま残す。
1月23日 月 小保方さん帰国初出勤。
1月24日 火 129xB6-GFPの赤ちゃんマウスを雌雄取り混ぜて数匹渡された。STAP作成開始。
1月31日 火 若山さんクローン胚-1移植
2月02日 木 若山さんクローン胚-2~8移植。
2月04日 土 3.5日クローン胚-1胚盤胞胚の取り出し、ntES細胞培養開始。
2月06日 月 3.5日クローン胚-2~8胚盤胞胚の取り出し、ntES細胞培養開始。
2月16日 木 ntES-1樹立
2月18日 土 ntES-2~8樹立
2月20日 月 ntES-1~8の2N・4Nキメラ胚移植。
2月24日 金 ntES-1~8の2N・4Nキメラ胚のインヴィトロ培養後、再度インナーセルマスを取り出しソートしてリシピエント細胞を排除して幹細胞FLS樹立とする。3月03日 土 ntES-1~8の2N・4Nキメラの12日胚の帝王切開。(Extended Data Figure7-d-下)3月11日 日 ntES-1~8の2N・4Nキメラの出産。(Figure 4-c,Extended Data Figure7-a-右、7-b)3月12日 月 西川氏本人主張TCR再構成アドバイス。(手記は4月のネイチャーリジェクト後)
3月22日 木 アニマルカルス命名。
4月06日 金 CDB若山研メンバーがFLSの4NキメラのDNA抽出。(出産から26日後に当たる)4月19日 木 129B6FES1(僕のマウスES)樹立培養開始。(桂報告)(若山さん持ち出しリストでは5/25)(1~6まである)
4月24日 火 米国特許仮出願。
4月27日 金 倫理委員会にて竹市所長の知るところとなる。(若山研究室から神戸研究所研究倫理第一委員会に「STAP 現象 をヒト体細胞に適用する計画」が申請され、小保方氏が説明 )
4月30日 月 ntES-1~8由来2N・4Nキメラメイティング。4月中にネイチャーリジェクト。
5月20日 日 ntES-1~8由来2N、4Nキメラ子出産。(2Nは兄妹交配 Extended Data Figure 8-i 小保方氏が抽出した DNAあり、4Nは戻し交配 Figure 8-j)(2Nは兄妹交配 Extended Data Figure7-c 小保方氏が抽出した DNAあり、4Nは戻し交配?結果無し)5月25日 金 CTS1 樹立培養開始。129B6F1ES(僕のマウスES)樹立培養開始。129B6F1TS(僕のマウスTS)樹立培養開始。(若山さんの持ち出しリスト)
6月06日 水 セル誌リジェクト。(TCR再構成の切り貼りレーンが使われている)
7月09日 月 CTS11~13樹立培養開始。
8月13日 月 AC129-1,2樹立培養開始。
8月21日 火 サイエンス誌リジェクト。
始めの仮定で、2011年度に作られた幹細胞をソートし直したものが既にあると仮定変更する。この場合は、FLSは作る過程を省略したところから始まればいいだけで、日程は変わらない。
小保方さんの論文作成に関して大事なのはSTAPキメラであって、FLSは幹細胞化実験なので、最初の三誌投稿段階ではFLSのキメラ実験は入ってない。笹井さん参加後の論文でFLSの実験が入れられることになるが。それは三誌リジェクトされた後、ネイチャー二報同時投稿の話になった時にヴァカンティが抵抗して、小保方さんは米国に帰った。結局小保方さんに渡したその時の若山さんのデータがベースになっている。
小保方さんの実験はキメラが出来なかったということで終わっている。若山さんのntES化実験は進められているが、小保方さんの論文はキメラは出来てないのにできたと騙されて書くという状態になっている。若山さんは通させないつもりだから構わない。ヴァカンティのティシュー誌にぶら下げたら終わり。後は自分のntES化実験を、小保方さんを山梨大での助手にしてから、本格的に研究して、今度は嘘でないntES化した結果として、ちゃんとした本物の論文を書かせる。その前準備をしている。
①小保方細胞核使用ntESの無限継代の証明。(細胞増殖率表の作成実験)
②小保方細胞核使用ntESのキメラ形成能の証明(2N・4Nキメラの作成実験)
③小保方細胞核使用ntESキメラの生殖能力確認(2N・4Nキメラのジャームライントランスミッション確認実験)
④小保方細胞核使用ntESの遺伝子解析
⑤小保方細胞核使用ntESの4Nキメラの体細胞DNAの採取。
⑥小保方細胞核使用ntESキメラの胎盤蛍光の免染確認。
⑦小保方細胞核使用ntESをFgf4誘導したFI-SCキメラの胎盤蛍光確認。
若山さんは所謂二兎を追ってる。
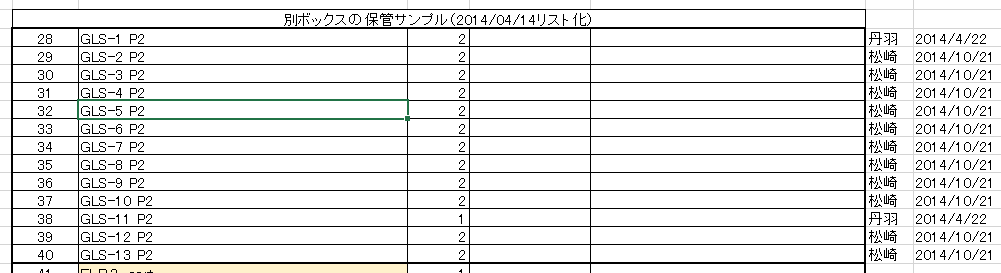
(FLBとは何か)上記日程の検討に入る前に若山さんの事後MTAの添付リストを再掲しておこう。

FLBの意味はリスト内に書かれているようにF1 Limphocyte Blastcystなのだから[F1の白血球胚盤胞」という意味である。分類はSTAP-SCであるが、奇怪なことに桂チームはこれを一切分析していない。
FLB-1~8とあって上から4,5,6行目がそれに該当する。1/31,2/2,2/3の三度に分けて作成準備されている。
1/31 2Nキメラ胚を7個使って2個樹立された
2/2 4Nキメラ胚を8個使って4個樹立された
2/3 2Nキメラ胚を3個使って2個樹立された
理研側に残されている試料は以下です。
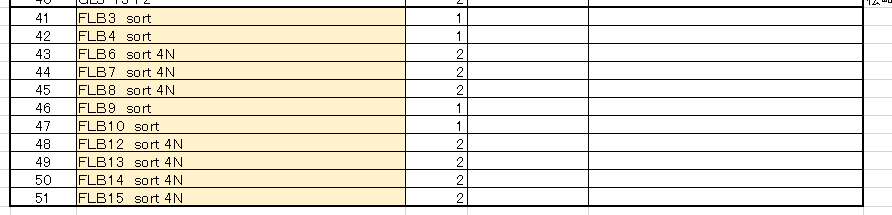
通しナンバーが違ってます。少なくとも15番まである。作成途中のものと考えてもうまく説明できませんね。一年前に持ち出したときのメモは有るはずですが、1年後に約束通り事後MTA締結した時にもちゃもちゃと弄っているんでしょうね。残していたものとのディスクレが出ている。
ここにあるsortという文字は細胞をGFPの有無でsortしているんですね。このFLBが何かということについては、岡部研究室のブログに書き込まれた若山さんのコメントから推測している。
>>
No. 2172 (2014/02/02 02:51) Cumulina
べさま コメントをありがとうございます。核移植を一番の専門にしているのに、核移植のいらない初期化方法を発表して、自分で自分の首を絞めている論文の関係者です。今回の小保方さんの発見はすごすぎたのかレフェリーに相手にしてもらえず、ずいぶん苦労しました。いまマスコミでリケジョとか違う方向で話題になっていますが、本当にすごい研究者で膨大な実験を徹夜続きで行いました。論文ですが、サプリにたくさんのデータが乗っていますが、それもほんの一部です。たとえば細胞の樹立がなかなかできず、STAP細胞を注入したキメラ胚を使って初めて樹立に成功したデータは、当初それだけで論文にするつもりでしっかりした表と解析を行っていたのですが、途中から直接簡単に樹立できるようになり、葬り去られました。実験中にどんどん発展していったのでしょうがないですが、STAP細胞の将来がすごく楽しみです。「STAP細胞を注入したキメラ胚を使って初めて樹立に成功したデータ」という言葉から幹細胞化の手法ではないかと推測した。このことは和モガさんも書いていたはずですね。彼は4Nをsortする意味を掴みかねていましたね。というのも4Nは胎生致死という知識があって、この早い段階ではまだ死んでないという可能性があるんですが、実証事実としてはど素人は知りませんからね。
我々がこのことに気づいたのは12/27のテラトーマからGFPの無い組織が出たという事実からです。桂チームは体細胞切り出しだと考えたのですが、ES捏造仮説なのに体細胞を切り出すはずが無いということにすら気づいていないんですから愚かと言われても仕方ありませんよね。まして、体細胞ならヌードマウスなんですから組織の遺伝子解析をしたらすぐわかることですが、それをせずにGFPがないから体細胞だと短絡した。愚かしいというより、結論ありきの犯意すら感じるところです。
我々の推定ではこのFLBはAcr-CAGだと思いますよ。若山さんはそれを今回のネイチャー論文とは別の自分の実験試料だから関係ないという説明で言い逃れているんじゃないでしょうか。若山さんを疑わないというのが桂チームの基本方針ですからそれを押しての調査はしないということですね。小保方さんに対しては病院にまで押しかけて行って、たまたまその場にあった実験ノートを心身不調の彼女に対して配慮もなく持ち帰りそのコピーを意図的にNHKに流出させるという公務員法違反犯罪まで犯しているんですから呆れる。
それはともかく、このFLBと同一の趣旨で作られているもう一つの細胞試料がありますね。若山さんの持ち出しリストの8行目のGLです。GOF Limphocyteです。GOFマウスの白血球のSTAP-SC分類の細胞ですから、蛍光細胞をCumulina書き込みの手法で作ったものです。我々のntES仮説ですと、GOFの酸浴蛍光細胞の核を使ったクローン胚からのntESです。若山さんはこの細胞を小保方さんに渡していない。しかし、後のリストにはその存在を書き込んでいる。Cumulina書き込みと対応しているアリバイ作りでしょうね。リストはCumulina書き込みの後に提出されているものです。FLBは理研側に残されていますからこれが何かは説明しないといけなくなる。Cumulina書き込みは表と解析があると言ってます。調査段階で押さえられるサンプルがあると、それを説明しなければならなくなる。FLB は残してしまっている。結果GLも残さざるを得なくなる。Cumulina書き込みというのは調査を予期した言い訳なんですね。細胞は既にntESになってしまっているものです。桂調査チームはGLを取り寄せて解析することも無論していない。性別を調べるだけでも分かることがあったでしょうね。今やもう事件は解決したとして廃棄してしまってるでしょうね。廃棄されていないのは理研側の試料だけでしょう。
GLがあるということはF1の幹細胞もないといけませんが、それは何も書かれていない。最初のキメラ成功時に幹細胞もできたと言ってますよね。我々は12/27テラトーマの上から注射されたのがそのF1のソート前の幹細胞だったとみています。だからリシピエントのインナーセルマス由来のESからできたGFPの無いテラトーマの切片が作られたんですね。それはどこかにあるに決まっていますが、リストに書かれていない。そもそもこの時のキメラすら残されていない。若山さんが廃棄しているんですね。かろうじて見つかったのが2011/11/28日付の4Nキメラの画像でしたね。小保方さんはその画像を2012年に入って胎盤が光ってるから調べてくれと頼まれたときの画像として論文に掲載していましたが、それは違うと若山さんが言いだしたがために、逆に2011年の写真の存在が明るみに出たのみならず、その時のマウス背景が「僕のマウス」ではないということまで知られてしまった。この写真が本当に小保方さんの写真選択の間違いかどうか、桂報告は確認したと言っているだけで、その写真を公開していない。テラトーマに関しては比較写真をつけているにも関わらず、こんな大事なPC内にあったという写真と論文の写真とどう同じなのかの比較写真はないのです。言うまでもありませんが、裁判ならこんな主張をしたら比較写真を出せと要求されます。出さなかったらその時点でこんな重大な事実に関して嘘をついていると判断されます。言ったことは事実証明しないと裁判官の心証を害して敗訴しますね。
STAPではできないからESでテラトーマを作ったのだと主張しておきながら、ESでできなかったから体細胞を切り出したなんてことを言ったらブーメランが自らの後頭部に帰ってくることくらい分からないのでしょうか。裁判官が素人だからと言ってその知性を舐めてるんでしょうかね。
(F1の幹細胞だけがなぜもう一度作られたのか)実験日程計算ではF1だけを書いていますが、無論GOFでの実験も同時に行われていることになっている。でも変なのはGOFマウスのキメラ胚を使った幹細胞化の実験は無いんですね。持ち出しリストの上から7行目です。
1/31に2STAP /wellを13作って、13ライン樹立、GLS1~13ですね。GOF Lymphocyte Spheresです。達成率100%ですね。小保方さんが作ったSTAP細胞をES培地に入れたら増殖したというんですね。これが小保方さんには全くできなかった。又丹羽さんの検証でもできなかった。小保方さんがリヴィズ実験中にできないと訴えても、若山さんは自分の手技だと言って教えなかった。桂チームはこれを学生のGOFntESによる捏造だと結論した。しかし小保方さんによる捏造なら若山さんは何も手技の必要なことはしてない理屈になる。山梨にSTAP細胞を持って来い。目の前で作ってやると言えばいいことだ。そうできないのはntES化しているからだ。我々はsortしたGLそのものだと考えている。
GOFのntES化実験はGLをソートしただけのもので実際には行われていないと考えられる。理研側の残存サンプルは以下である。
既述しているように丹羽さんの持ち出しは性別の問題があって核型解析に出したときのものですね。これは学生のGOFntESと同じものと確認された。当時ラボを離れていた学生本人が自分の作ったGOFntESをどこかにまだ持っていたのか、それを理研に提出したのかどうかは何も報告されていない。が、小保方さんが学生から貰ったと言われる細胞はあった。以下の分である。

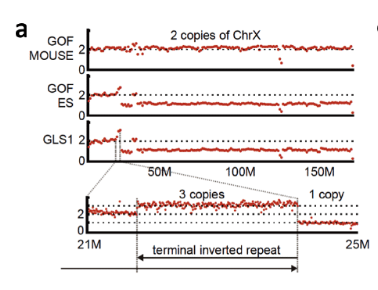
GLS1=GOF ESだということは桂チームが遺伝子異常の存在で証明している。BCA報告の図を以下に添付しましょう。
小保方さんが犯人でなければ試料の中身の入れ替えが逆に証明されたことになりますよね。今Ooboeさんのパートナー氏が検察に資料提出されていますね。ここは我々が早くから小保方さんか若山さんのどちらかが犯人だということにしかならない証拠なのだと主張していたものです。小保方さん擁護派の人でもどちらも犯人でないという人たちが多かったですね。中身が入れ替えられてなかったら小保方さんが犯人です。若山さんが犯人ならこの中身は入れ替えられている。だからX染色体上の逆位接続が一致してるんです。若山さんが本当に小保方さんから貰ったGOFのSTAP細胞をそのまま培地誘導で幹細胞化したのなら小保方さんにプロトコルを教えるだけで簡単にできたでしょうよ。小保方さんができないと言ったらウソつくなと言わないといけない。誰にでもできる。すぐSTAP細胞を持って山梨に来いと言えばいいだけです。3日もいれば増殖を確認できる。
桂チームは目の前に誰が犯人であるかの証拠を発見していますね。小保方さんが犯人だと思うなら、このサンプルの中身を若山さんが入れ替え得なかったという証拠を調べたらよかったんです。その証拠が出たら小保方さん犯人確定です。逆に若山さんが犯人ではないのかと疑うのなら、中身を入れ替えているということを証明したらいいんです。実は調査サンプルは全部若山さんが自分の行ったことを隠すために都合よく中身が操作されているものですから、このGLSだけではないので、どれであってもサンプルの提出経路の説明に嘘があったら若山さん犯人が確定するんです。今Ooboeさんのパートナー氏が検察に調査依頼しているのがその件なんですね。
細胞の大きさは言わずもがな若山さんの説明には他にたくさんの矛盾があって、嘘の傍証にことかかない。中身が入れ替えられていないと仮定すると他の矛盾が説明できなくなります。あちこち押しピンで止められていますから一か所外したからと言って身動きできないんですね。ということは外してみる仮定も間違いだと分かる。事件はそういう論理構造になっているんですね。
(GLSに関するDORA氏の調査結果)このGLSに関してはそもそも雌雄の問題があって、旧DORAブログに詳しい調査結果があります。何れ見れなくなる恐れがあるので、まずはここに保存しておきましょう。
GLSの性別に関する解析データ。
2017/4/23(日) 午後 2:39 日記 練習用
久々にSTAP関連なんですけど。ずっと以前、GLSの性別について、触れたことがありまして、だいたい以下のような経緯です。
2014年6月16日の記者会見で、若山氏は「GLSの13株すべてがメス」と発表した。しかし、そのあと、若山氏はGLSの性別について、CDBの発表との間に齟齬が生じていることに気づき、訂正している。『6月16日に山梨大で行った会見内容の一部修正、およびNatureに掲載された撤回理由書の訂正について』(7月22日)において、若山氏の方から次のように訂正した。
会見時に、もう一種類のSTAP幹細胞であるGLS(Oct4-GFP遺伝子をもつB6由来とされる)は細胞株の1番から13番まですべてメスの細胞だと報告しました。しかし、CDBとともに詳しく精査したところ、GFPすべての細胞株はオスだということがわかりました。原因は、GLSのY染色体は性決定遺伝子SRYを含むY染色体の重要な遺伝子群に欠損があり、用いたプライマーではY染色体が存在しないと誤判定されてしまったからです。
ところが、このあと、桂調査委の報告書では、再びオスからメスへと性別が逆転している。若山氏が「CDBと詳しく精査した」と述べているにもかかわらず、それをあっさり覆している。しかも、そのことに対する桂調査委からの説明はいっさいなかった。そこで、「CDBとともに詳しく精査した」という、その解析データを理研の情報公開窓口を通じて入手したところ、次のようなものであった。まずは核型解析についてである。
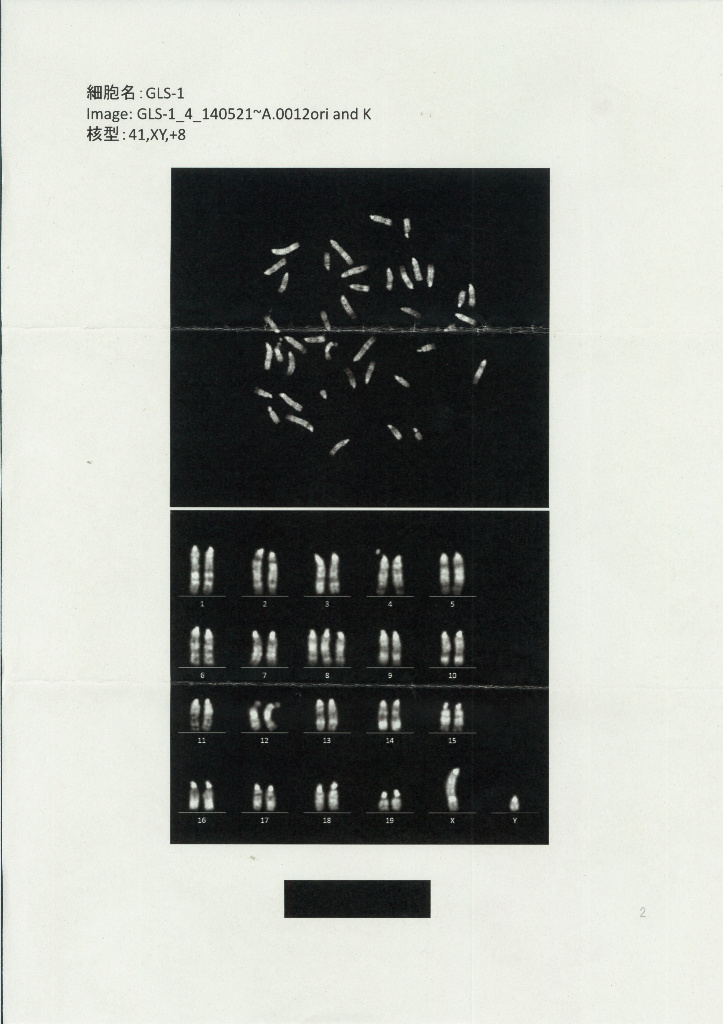
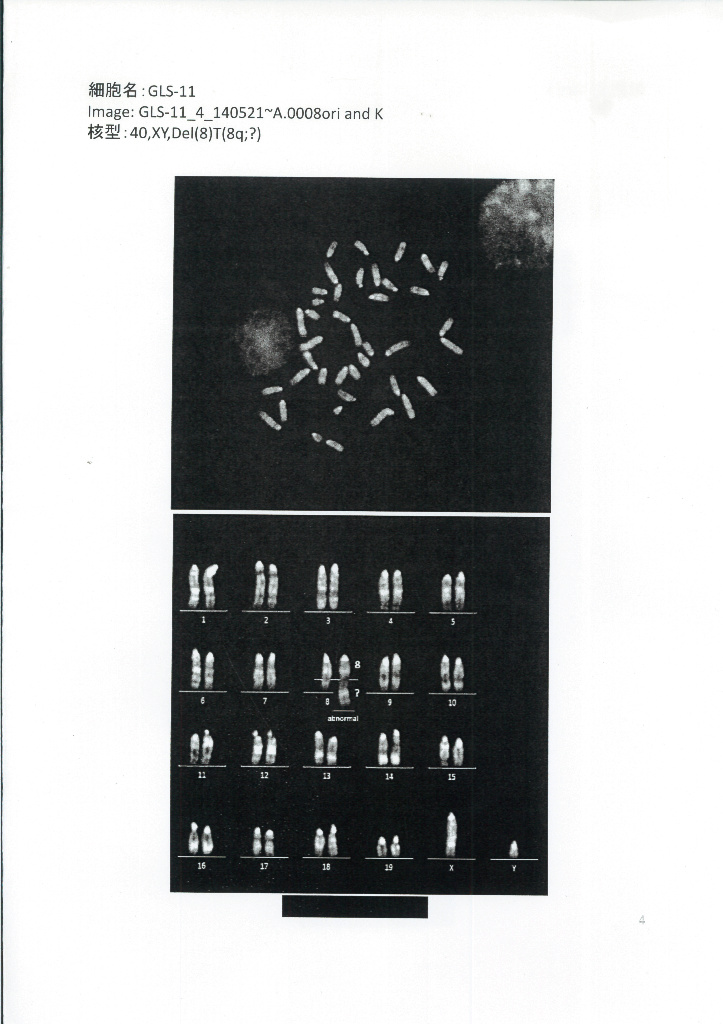 以上のように、核型解析の専門機関から丹羽仁史氏に対し、GLS-1とGLS-11は、ともにオスであると報告されているようです。GLSの性別に関する解析データ(その2)。
以上のように、核型解析の専門機関から丹羽仁史氏に対し、GLS-1とGLS-11は、ともにオスであると報告されているようです。GLSの性別に関する解析データ(その2)。
2017/4/24(月) 午前 10:59 日記 練習用
次に、PCRによる解析データです。以下の通りです。
 いちおう、説明部分を書き出しておきます。
いちおう、説明部分を書き出しておきます。
この結果、Eif2s3yとSsty2のPCRは、B6♂で予想通りのサイズのバンドを検出し、129♀では全く検出しない事から、Y染色体上の遺伝子を特異的に検出していると考えられた。これに対し、予備的検査で用いたZfy1ならびに新たに設計したSryのPCRは、B6♂で予想通りのサイズのバンドを検出するものの、129♀でも薄いバンドが見える事から、特異性が低いと考えられた。
FLS3とAC129-1は共にSsty2,Eif2s3yが検出され、雄であると判定された。これに対し、GLS1,GLS11はSsty2は検出されるもののEif2s3yは検出されず、Zfy1のバンドも129♀同様に薄かった。これらの結果と簡易核型解析の結果をあわせて考えると、GLS1,GLS11は部分的に欠失したY染色体を有する雄の細胞であると考えられる。
つまり、このときはオスに特異的なSsty2遺伝子の検出と簡易核型解析との結果によって、Y染色体が部分的に欠失したオスだと判定されたようです。GLSの性別に関する解析データ(その3)。
2017/4/25(火) 午前 8:55 日記 練習用
桂調査委の調査報告では、GLSについて「大きな欠失+末端重複逆位接続」という構造異常を指摘していた。で、「末端重複逆位接続」についてはスライドで説明されていたのだが、「大きな欠失」についてはデータとして示されないので、それがどの程度の欠失なのかいまひとつわからなかった。そこで、情報公開窓口を通じてそのデータを入手したところ、次のようなものであった。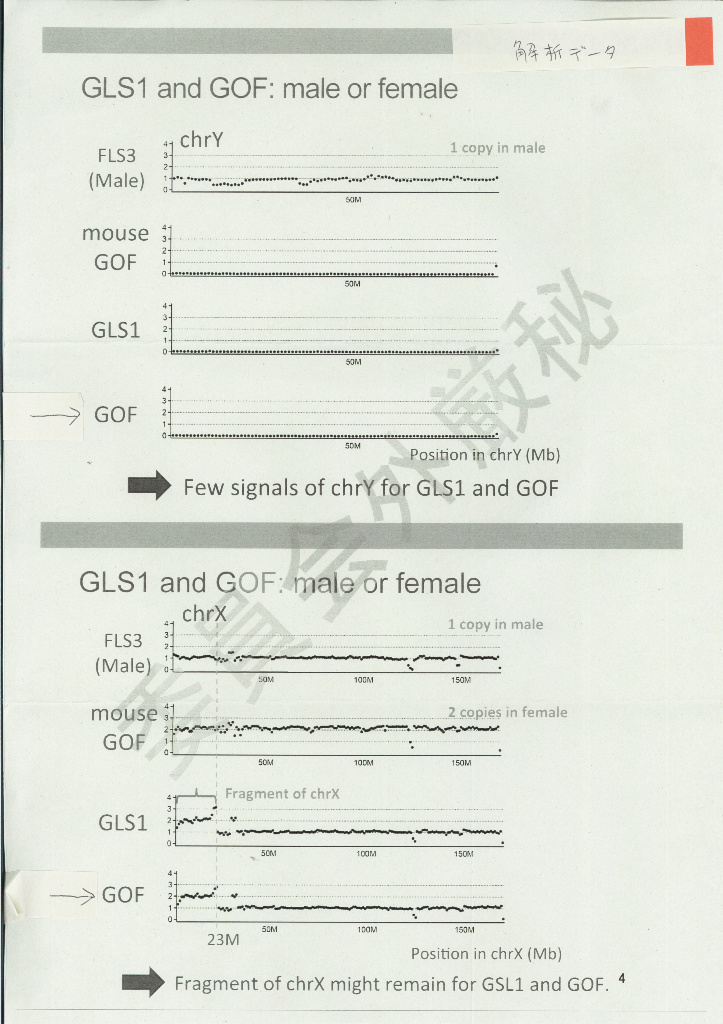 このデータは後に『STAP cells are derived from ES Cellls』には掲載されたようなのだが、なぜか桂調査委の報告書には載らなかった。これをみると、およそ170Mのうち23Mしか残っていなかったらしい。となると、核型解析でY染色体だと断定したものは、じつは非常に大きく欠失したX染色体だったと考えることができるわけだ。
このデータは後に『STAP cells are derived from ES Cellls』には掲載されたようなのだが、なぜか桂調査委の報告書には載らなかった。これをみると、およそ170Mのうち23Mしか残っていなかったらしい。となると、核型解析でY染色体だと断定したものは、じつは非常に大きく欠失したX染色体だったと考えることができるわけだ。
しかし、まあいちおう、核型解析の専門家がみたらどう判断するのか、念のために、核型解析を専門にしている会社に電話して、「ある核型解析を判定してもらえないか」と頼んだところ、そこの研究者が快く承諾してくれた。そこで (先入観のないよう細胞の正体を伏せて)見せたところ、一瞥して躊躇なく「これはオスです」と断言した。これにはむしろこちらが驚いて、「それじゃあ大変なことになる。これはゲノム解析ではメスなんです、欠失したX染色体なんです」というと、ちょっと困惑して「それならもっと詳しく調べてみないとわからない」という答えだった。
で、まあ、核型解析は専門家の勘違いだったとしても、それならSsty2遺伝子の件はどうなるのか。それが気になった。GLSの性別に関する解析データ(その4)。
2017/4/26(水) 午後 2:33 日記 練習用
で、それがメスであることと、Ssty2を持っていることと、どのように両立するのか、もしかすると、なにか納得のいく説明でもあるのかな、と、このことを気まぐれ先生にうかがってみたのですが、正直、先生も困惑気味でいらっしゃったようで、「説明できない」とのお答えでした。(なお、気まぐれ先生は解析にはいっさい関わっていらっしゃらなかったとのことです。)
「遺伝子をいじくった可能性もある」とのことでしたが、そういう可能性を考えない限り説明できないということだと思われます。
また、ES細胞の樹立時に、このような大きな欠失が生じることがあるのでしょうか、とお尋ねしたところ、次のようなお答えでした。
「普通メスの体細胞では 、X chromosome inactivation ということが起こっていて、2本の染色体の中のどちらかが不活性化され、発現しないようになっています。この現象が起こるのがES細胞がとられる時期の発生段階です。2つの活性なX染色体があることは、ES細胞にとっても都合が悪いようで、2つの活性なX染色体をもつES細胞が得られることは稀で、どちらか一方が失われる、あるいは、大きな欠失を起こす、また、得られたとしても培養している間に、このようなことが起こるのは普通です」
この説明ですと、X染色体に大きな欠失があったことは、それがES細胞であることの有力な証拠だったことになります。しかし、そうなると、桂調査委が報告の際に「末端重複逆位接続」のデータだけを示し、大きな欠失のデータを示さなかったことはむしろ不自然に思えます。
結局、確実な結論は出せなかったのですが、どちらかというと、データを全て公開せず、詳しい説明もないまま公的に確実な結論を出す方が軽率であるように思われます。DORAさんの考察はここで終了しています。佐藤さんはこの問題を2015/12/7発行の『STAP細胞 残された謎』で触れています。後の2016/2/14に発行した続編『STAP細胞 事件の真相』ではこの問題に触れていません。DORAさんが『GLSの性別に関する解析データ』のその1~4を書いたのは2017/4/23(日) 午後 2:39から2017/4/26(水) 午後 2:33の間です。佐藤氏はDORAさんではないかと言ってる人もいますが、個人的なことには興味がありません。ただこの件に関する書き出しは佐藤本と同じです。和モガさんも無論この問題には触れています。私は最初和モガさんが佐藤さんなのかなと思っていて佐藤本第三弾を期待しましたが、彼は若山さんによる成果の隠蔽という推測ですが、DORAさんはただ真相究明姿勢ですね。どちらかというと後者だったかもしれませんね。まあ、どうせ皆匿名でやってるんですからどうでもいいことです。
しかし、扱われている問題はどうでもいいというわけにはいきません。いまだに放り出されたままになっている。若山さんは2014/3/10の共著者に対する論文撤回要請の後に放医研にサンプル解析を依頼しています。若山さんの記者会見は2014/6/16でしたが、この記者会見の席上で山梨の若山研でGLS1~13の性別を確認してすべてメスであったと発表した。一方、丹羽さんの記者会見は若山さんより2か月以上早く、2014/4/8に行われましたが、その2週間ほど後の2014/4/22に理研側に残されていたサンプルを持ち出しています。その解析結果はDORAさん取り寄せ資料にある通り2014/5/27です。数日後には丹羽さんは結果を受け取っているはずですので、すぐに伝えたら2014/6/16の若山さんの記者会見ではすべてオスであったと発表されたはずですが、どうも連絡のずれがあるようですね。若山さんは自分で調べた通りにメスだと発表した。そして記者会見の後の、2014/7/22に、『6月16日に山梨大で行った会見内容の一部修正、およびNatureに掲載された撤回理由書の訂正について』(7月22日)を公表した。DORAさんのブログからそれを再掲すると以下です。
>>
会見時に、もう一種類のSTAP幹細胞であるGLS(Oct4-GFP遺伝子をもつB6由来とされる)は細胞株の1番から13番まですべてメスの細胞だと報告しました。しかし、CDBとともに詳しく精査したところ、GFP(ママ。GLSのタイプミスだと思われる。)すべての細胞株はオスだということがわかりました。原因は、GLSのY染色体は性決定遺伝子SRYを含むY染色体の重要な遺伝子群に欠損があり、用いたプライマーではY染色体が存在しないと誤判定されてしまったからです。これを見ると、丹羽さんが結果を受け取ってから検討があって、若山さんに伝えられたのが遅れたのだと推測されるところです。ここで若山さんはY染色体の性決遺伝子をSRYだと考えていて、それを検出するプライマーでPCRに掛けたのだが、それがなかったからメスだと判断したのだということになる。
ところが、丹羽さんは自分で解析に出したGLSの性別に関して意見を書いている。核型解析の結果はオスでした。写真を誰が見てもオスです。ところが、丹羽さんの報告に曰く、これは山梨大と理研とで行った予備検査の結果であるメスという結論と矛盾していたと。山梨大と理研で予備調査をして、同じ結果だった。メスだったということです。そして、核型解析結果を見た後に、丹羽さんがもう一度PCR検証をやり直した。そして上の表にあるような結果を得たんですね。以下に整理します。数字は遺伝子座の開始位置番号です。上から順番に並んでいるものです
>>
725207 Zfy1(丹羽さんの予備調査ターゲット遺伝子)
B6♂〇 129♀△ FLS3〇 AC129-1〇 GLS1△ GLS11△
1010543 Eif2f3y(丹羽さんが新たに設計したターゲット遺伝子)
B6♂〇 129♀✖ FLS3〇 AC129-1〇 GLS1✖ GLS11✖
2662471 Sry(若山さんの予備調査ターゲット遺伝子、丹羽さんは今回初めて設計)
B6♂〇 129♀〇 FLS3〇 AC129-1〇 GLS1△ GLS11△
67045883 Ssty2(丹羽さんが新たに設計したターゲット遺伝子)
B6♂〇 129♀✖ FLS3〇 AC129-1〇 GLS1〇 GLS11〇
△はありですが、バンドの出方が薄いものです。薄いですが他に紛らわしいものがありませんから有りは有りなんですね。人によって見えた方が違うということも無いと思いますがね。確認してもらえばいいですね。問題は肝心のSRYですね。129♀〇ではY染色体特異的遺伝子では無いではありませんか。まずもってど素人には理解できませんね。素人常識ではメスにY遺伝子特異的遺伝子のバンドがあったら、それはメスと言いながらに実はオスであったか、SRYが実はY遺伝子特異的遺伝子では無いかのどちらかです。
そこにもってきて、丹羽さんはこんなことを言ってる。
>>
この結果、Eif2s3yとSsty2のPCRは、B6♂で予想通りのサイズのバンドを検出し、129♀では全く検出しない事から、Y染色体上の遺伝子を特異的に検出していると考えられた。これに対し、予備的検査で用いたZfy1ならびに新たに設計したSryのPCRは、B6♂で予想通りのサイズのバンドを検出するものの、129♀でも薄いバンドが見える事から、特異性が低いと考えられた。あのねえ。冗談も休み休みお願いしたいものですよ。ど素人でも趣味で科学ファンというのは居て、ブルーバックスシリーズ程度なら楽しくて全部読んだなんて人もあるよねえ。でもこんな論理の運びは読んだことないでしょ。今Y染色体特異的遺伝子を探してるんでしょ。そしてその遺伝子座の位置は分かっていて、最大26桁、つまり4の26乗に一つしかないという位置を特定してプライマーを設計している。4の26乗って全遺伝子座数より大きい数値だからね。そしたらメスにもあったからこれはY染色体特異的遺伝子では無かったようだって。。。
変でしょ。何を言ってんの。
若山さんは「GLSのY染色体は性決定遺伝子SRYを含むY染色体の重要な遺伝子群に欠損があり、用いたプライマーではY染色体が存在しないと誤判定されてしまった」と言ってるんですよ。SRYはY染色体の重要な遺伝子だと思ってGLSの性を決定したんでしょ。欠失があっての誤認はともかくとして、丹羽さんは「新たに設計したSryのPCRは、B6♂で予想通りのサイズのバンドを検出するものの、129♀でも薄いバンドが見える事から、特異性が低いと考えられた」とおっしゃった。
これが何を意味しているかというと、新たに設計したSryのプライマーがいけなかったと言ってて、SRY自体がY染色体特異的遺伝子では無かったという意味ではないはずだ。でも挟んだプライマーは二つのそこしかないという絶対的位置です。
GPSで場所が特定されて、正確にカダフィの愛人の家の煙突の四角い穴からクルージングミサイルは突入してきた。そういう時代です。
二つの点の間にSryなるY染色体特異的遺伝子がある。挟み方が悪いとX染色体を含む他の染色体のどこかにSryなるY染色体特異的遺伝子の無い断片があり得ると。
変でしょ。SryはY染色体特異的遺伝子では無かったという意味にしかならないよね。
細胞生物学ってその程度の学問なのか。未だにY染色体上にある特異的遺伝子すら特定されていないのか。こんな調べればわかるようなことすらまだ試行錯誤中だとしたら、ここに従事している学者たちって仕事してるの? 税金で只飯食らってるゴクツブシかね。マスコミの羽織ゴロツキと同じレヴェルなの? 無論、嫌味で言ってるんだけどね。
若山さんはSryはY染色体特異的遺伝子だと言った。丹羽さんは違うよと言った。若山さんの言ってることが間違ってたとしたら、若山さんは勉強不足のアホなのかね。それとも嘘つきなのか? 嘘だとしたら、オスであったら困ることがあったのかな。
予備調査で丹羽さんも雌だと思っていた。ただし、Sryではなくて丹羽さんはZfy1で調べたと書いていますね。再検査の結果は以下でした。前回がどうであったかは分からないが、今回は△を無いと判断するメスになりますね。
>>
725207 Zfy1(丹羽さんの予備調査ターゲット遺伝子)
B6♂〇 129♀△ FLS3〇 AC129-1〇 GLS1△ GLS11△
丹羽さんもメスだと予想していた。しかし、核型解析結果がオスだったので、新たに調べ直して、結局、予備調査で自分の使ったマーカーと若山さんの使ったマーカーは捨てて、新たに設計したEif2f3yとSsty2をY染色体特異的遺伝子として使うことにした。
>>
1010543 Eif2f3y(丹羽さんが新たに設計したターゲット遺伝子)
B6♂〇 129♀✖ FLS3〇 AC129-1〇 GLS1✖ GLS11✖
67045883 Ssty2(丹羽さんが新たに設計したターゲット遺伝子)
B6♂〇 129♀✖ FLS3〇 AC129-1〇 GLS1〇 GLS11〇
どちらもB6♂〇 129♀✖ですね。これぞY染色体特異的遺伝子ではありませんか。そしてSsty2があるのですからオスは決定で、 Eif2f3yが無いから欠失していると結論した。当然ですね。
当然でないのは、そもそもお前たちの言ってるY染色体特異的遺伝子というのは既存知識だったのかという疑義です。今度は、そんな馬鹿なことはない。Y染色体特異的遺伝子というのは既存知識だ、という前提で考えなおしてみましょう。専門家はどちらか知ってますからね。ど素人はどちらか分からないときは両方とも考えるだけです。専門家が正直だったら不要な手間ですけどね。専門家が嘘ついてるような低レヴェルの未開社会だから仕方ない。
(コントロール)コントロールはB6♂と129♀です。Y染色体特異的遺伝子はZfy1,Sry,Eif2s3y,Ssty2です。ところが前者2つには129♀にも薄くバンドが出ている。この時点で既に丹羽さんは説明する義務がある。
Zfy1,Sry,Eif2s3y,Ssty2はすべてY染色体特異的遺伝子でしょということです。129♀にバンドが出てはいけませんよね。まずこの説明が必要です。これらの遺伝子はX染色体上には無いからY染色体特異的遺伝子だとされているはずです。その機能に関してはまだ研究途上であっても構いませんが、今調べているのはY染色体の有無です。X染色体上にも同じものがあるような遺伝子を調べることはあり得ませんね。マウスの全ゲノムは既に読まれていますよね。同じ配列がX染色体上に無いからこれらの4つが選ばれているはずです。
なぜ、Zfy1とSryが129♀にも薄く出ているのかの説明が必要です。129♀に129♂の細胞が少し入ったのではないのか。或いはそれが考えにくければ例えばこの細胞が血液だとして、B♂の採血の残りが少し入ってしまったのではないか。でもその場合、Eif2s3yとSsty2でも同様の薄いバンドが出るはずです。とても理解しにくい現象です。
一方若山さんの説明は丹羽さんとは違っている。SRYを性決定遺伝子であると考えているらしいことが書かれている。以下です。
>>
原因は、GLSのY染色体は性決定遺伝子SRYを含むY染色体の重要な遺伝子群に欠損があり、用いたプライマーではY染色体が存在しないと誤判定されてしまったからです。明確でないが、若山さんはマーカーとしてSryを使った。で、自分が山梨大に持っていたGLS1~13の全株を、SryがY染色体特異的遺伝子であると思って、PCRで確認したら全株に無かった。だから全株メスと言った。メスだと思っていたが、丹羽さんオスだと言ってる。丹羽さんが調べたんだから間違いないだろう。ではY染色体特異的遺伝子であるSryの遺伝子座を欠損しているのだなと理解した。結論だけは丹羽さんの結論に合わせた。
対して丹羽さんは若山さんと違って、Y染色体特異的遺伝子としてZfy1を選んだようである。そして理研側に残されていたGLS1と11(木星リスト28と38番)を予備的調査段階でPCRにかけたらどちらにも無かったから若山さん同様に全株メスだと結論していた。ところが別の機関に同じ細胞株を核型解析に出したところ、どちらも短いY染色体が見つかってオスだと結論された。
丹羽さんはなぜだと思ってもう一度PCRを本気でやり直したのである。まず今度はB6の雄と129の雌の細胞をコントロールにした。そしてY染色体特異的遺伝子を増やし。前述の4種とした。4つとも今でもY染色体特異的遺伝子とされているものである。
ところがとんでもない結果が出た。まずコントロール実験によって、自分の使ったY染色体特異的遺伝子だと思っていたZfy1と若山さんの使ったマーカーであるSryは129のメスからも検出されてしまった。
丹羽さんはまたも新発見をした。つまり、Zfy1とSryはX染色体にも存在しているということである。さもなければこのコントロール実験は失敗していることになる。実験が正しければ新発見なのである。Y染色体におけるZfy1、2に関しては精子の働きをコントロールしていることが分かったという論文が書かれていて、それはそれでいいのだが、その論文の紹介ではZfy1、2はY染色体にしか無いと"されている"という具合に紹介されているが、X染色体には無いと断言されてない。つまり丹羽さんのように実験で確かめられていないということになる。今回、雌からも薄く出ているので、これが実験の失敗でなければ新発見ということになる。この程度のことすら誰も確認してなかったなんて。だからこの分野の科学者たちはゴクツブシなのかと問うているのである。
Zfy1、2はオスの精子の働きをコントロールしている遺伝子だということは分かったが、X染色体上にもあるということになった。ただし、薄く出たということが何を意味することになるか。オスはXYです。メスはXXでZfy1はXにもYにもあるのですからどちらも同じ濃さで出ないとおかしい。となるとXYはどちらも働いていますが、XXの片方は相沢さんの言うようにX chromosome inactivationの所為だと考えると量的に半分しか出ない理由は説明できる。しかし、PCRに掛けるときは全部解除されてしまうのではないか。分からないことだらけです。
ただし、丹羽さんの説明には一つだけ疑念がありますね。最初の予備的調査ではZfy1が無かったから、若山さんと同じメスだという結論になっていたはずです。でも核型解析結果が出た後に再検査したときは薄く出たんでしょ。結果が違っている。
その齟齬を置いておいて、丹羽さんは取り敢えず、Zfy1とSryは129の雌からも検出されているのだからY染色体特異的遺伝子ではないとした。彼の言い方だと特異性が薄いとした。でも我々の理解では特異性はあるか無いかのどちらかですから、Y染色体特異的遺伝子では無いと言ってるのと同じです。そしてEif2s3yとSsty2はコントロールによってオスには出て、メスには出てないからY染色体特異的遺伝子だとした。そして、この二つのマーカー遺伝子のPCR結果によれば、GLS1と11にはSsty2が出ているからオスであるとした。ところが同じくY染色体特異的遺伝子であるはずのEif2s3yは出ていない。従ってこれはY遺伝子に一部欠失があってEif2s3yの遺伝子座を欠失しているのだと説明した。若山さんの理解とは違っていますね。どちらも合わせるとSryとEif2s3yの両方の遺伝子座を欠失しているオスという認識で説明できる。なぜ欠失しているにも関わらずオスなのか。Ssty2が検出されているからですね。
これを桂報告とBCA報告はメスだと逆転させたのである。DORA氏は「それがメスであることと、Ssty2を持っていることと、どのように両立するのか」と疑義を書き留めましたよね。
論理学の問題なら、Ssty2がY染色体特異的遺伝子でないか、GLSがメスだとは限らないかのどちらかです。アーティクル論文に載っている Extended Data Figure 8-b の核型解析写真です。
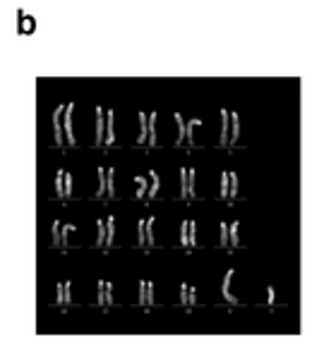
トリソミーはどこにもありませんね。しかも、この図のリジェンドは以下です。
>>
b, Q-band analysis (n = 4; all cell lines showed the normal karyotype). 参考にタカラバイオの宣伝広告を貼り付けて置きましょう。右側がマウスのESのオスのQ-band核型解析です。
 (並行する実験と三誌論文に採用されたデータ)
(並行する実験と三誌論文に採用されたデータ)小保方さんの最初のネイチャー論文の主旨はとてもシンプルなものです。自分が酸浴させた細胞をキメラ胚に入れたらキメラができたから自分の作った細胞は多能性細胞であるという主張です。4月に提出されて4月中にリジェクトされていますから、ジャームライントランスミッション実験結果は入れられていません。上記しているどちらの日程表によっても明らかですね。
我々のntES仮説では、そもそもスタンダードなプロトコルでキメラができたというのは嘘です。小保方さんを引き留めて、ヴァカンティへの約束である論文を出すというところまで行なって、最終的にはヴァカンティの主催誌であるティシュー誌にぶら下げさせる。そして山梨大に小保方さんを助手として連れて行くという目的のために嘘の論文を書かせている。
この嘘の論文は専門的にはバカバカしいほど嘘っぽく、かつ事実嘘なのでアクセプトされる気遣いの無いものです。そして、このことはこの時点で世界中で基本若山さんと小保方さんとヴァカンティの3人しか関与していない話だったんです。この時点で若山さんのついている嘘って他愛ないものですね。この論文を査読させられた人も多忙な中をいい面の皮だと思いますが、この最初の論文の査読は公表されてないのでわかりません。しかし三誌最後のサイエンスの査読は事件化したために例外的に現在サイエンス誌編集部によって公表されていますね。査読ではまあ、ESのコンタミだよと言われていて、どんな杜撰な管理しているラボなんだとけなされている。そしてこの査読者は正しいわけです。ntESもESなんですからキメラはESで作られている。別に杜撰な管理によるコンタミなんじゃなくて意図的に作っている別の実験結果を嘘と知らされずに著者が書いているだけだ。若山さんはそういう査読結果になるだろうなあと安心して書かせているんですね。採用されたら逆に困りますよね。でも絶対リジェクトされると確信している。自分が査読者でもリジェクトするでしょう。若山さんもネイチャーに何本も論文掲載されていますからね。そのレヴェルは知ってる。ヴァカンティもヴァカンティじゃないか。こんなんでキメラができるわけないだろう。再現されない限り認められないんだから自分の雑誌にぶら下げておいて、後で自分で再現するなり、人が再現したら第一発見者の栄誉が得られるじゃないか。再現できなかったら忘れられていくだけでしょう。そんな論文はこの分野に無数にあって、常態ではないか。自分にどうしてキメラができたのかなと問うて来たら、何かESのコンタミ事故でもしてしまったかなと言ったらおしまいじゃないか。この時点で何か事件の臭いがしますかね。しょうもない話だと思いますよ。
(幹細胞化実験)それに対して、若山さんの幹細胞化実験は真剣なものです。歴史的視点の無い人の陥る錯覚は、この時点でGOFマウスのOct4-GFPが大量に光ったという事実に関して、若山さんがこの細胞は何物かではあると信じ込んでいることを、後の知識から忘れることです。
事件化してからはむしろ自家蛍光の誤認は若山さんの言い訳にとって好都合だったのでノフラー氏や関氏の人脈を使って大々的に宣伝しましたが、自家蛍光を確認しない専門家なんてあるわけがないじゃないですか。自家蛍光なんて常時あるに決まっている。だからフィルター切り替えで確認するし、自家蛍光の継続時間が6時間程度ということを利用してライブセルイメージングで7日間の撮影も行っている。一コマ30分とか15分のインターバルで撮影すると6時間は12コマから24コマです。アーティクル添付の動画だと1週間分を38秒と97秒で早回ししている。1秒間に5コマから16コマ進む。つまり自家蛍光は1秒から2秒の間で発生したり消えたりするんです。それも最初のころに集中していますね。その後にずっと蛍光し続けているのは自家蛍光ではないんです。この実験は笹井研で行う以前に若山研で既に先に行われている。手記を読まない人には分からないことですね。
笹井さんは記者会見でこの細胞は何物かではあるといいました。多分若山さんが別の場所でそう言ってますから、若山さんに感化された言葉だと思います。若山さんは当時このGFP蛍光をキメラは出来ないまでも何物かではあると信じ込んでいたんです。これが小保方さんのこの細胞の核を使って自分のntES化技術を組み合わせれば何かしら新種の多能性細胞ができるのではないかと発想した直接の原因です。この若山さんの幹細胞化実験は何ら捏造などというものではありません。歴史的視点を得るための訓練の無い人たちには、笹井さん参加後のSTAP論文の趣旨に対して、ntESでキメラを作ったと言うと、時間的経過とその間の経緯の情報が欠落しているのでそんな馬鹿なことをするわけが無いと短絡するんですね。今回の事件で最も反省させないといけないのはマスコミのレヴェル低下でしょうね。単に頭が悪いと言う印象ですね。業界の人材の質が落ちてるようですね。斜陽化産業なのでしょうかね。
若山さんの実験は小保方さんの最初の論文と切り分けて考えるととてもまじめな研究です。これはこれで科学的な興味の尽きないとても面白いものだということは、キメラができたと嘘をつかれる前の小保方細胞が何であるのかという研究が若山さんを離れてヴァカンティ研で行われていたらどんなにか興味深い研究であったろうかということと同じですね。
でも事実はそうならなかった。それが事件の原因でオホホポエムの言ってることはある意味正しい。
(小保方さんの発見した現象)細胞は刺激を契機にリプログラムされ得るという発見はひょっとしたら小保方さんが世界で最初に発見した事実かもしれません。小保方さんがティシュー論文と博論を書き上げた後にヴァカンティと大和のアドヴァイスで、これは既存幹細胞ではなくて、トリチュレーションの物理刺激によってリプログラムが起きているのかもしれないという方向に研究仮説を変更したのが、2010年の12月のフロリダ会議でした。彼女は博士号取得後にヴァカンティ研のポスドクになる予定でしたから、その研究は米国で行われることになっていた。小保方さんは今までヴァカンティの胞子様細胞を見つけたのだと思っていましたが、最初からあるのではなくてできてきていた細胞を見つけて三胚葉分化させることに成功していたわけです。これが本当にできてきている細胞だったら世界で最初の発見です。
小保方さんの細胞は組織を選びません。どんな組織細胞からもとても少ない確率ですが、三胚葉に分化する細胞を発見している。ただ、これが本当にできてきているのか、或は各組織に存在している既存の幹細胞を拾い出してしまっているのかはまだ厳密には明確になっていない。それが今後の課題で彼女が米国で研究しようとしていたことです。
それに対して、全ての組織ではなく、筋肉という限定された組織細胞が刺激でリプログラムするという発見は、2011年2月に既にムーさんによって発表された。筋肉の損傷が速やかに修復されるという臨床事実は以前から知られていて、最初から存在している筋肉幹細胞が働いているのだと思われていた。でも、ムーさんはクレ追跡システムを応用して実はその筋肉幹細胞は最初から存在していたのではなくて、すでに筋肉細胞に分化してしまっている細胞が一旦多能性細胞にリプログラムされて、その多能性細胞から幹細胞ができるのだと証明した。筋肉というのは単核細胞が融合して二核の細胞になっているんですが、その二核の細胞が組織の裂傷を契機に単核に戻り、更に多能性細胞に一旦戻ってから、幹細胞を作るのだということを発見した。この発見が小保方さんの発見と同じ系統上にある研究だということは分かりますね。細胞は刺激を契機にリプログラムされ得るんだということです。
STAP騒動が小保方さんの辞任で終わり、今度は博士号の再取得指導を受けていた最中、2015年6月に、ムーさんの弟子のキンガ・ヴォイニーツがSTAP論文にヒントを得たのでしょうが、ムーさんの細胞がどの程度のリプログラム段階にある細胞なのかということを確認した実験結果を発表した。試験管内分化、テラトーマ形成、キメラ樹立が確認された。ジャームライントランスミッションだけが無かったのである。 ムーさんの発見したinjury induced muscle-derived stem cell-like cells (iMuSCs)は多能性細胞だったのである。つまり筋肉になる段階の手前で止まっているものでなく、もっと以前の段階までリプログラムされているということである。この論文でキンガは小保方さんのティシュー論文を先行論文の一つとして引用している。
(若山さんが発見したと思った現象)若山さんは小保方さんの細胞核を使用してntESを作ってみようと考えた。この時に大きな勘違いがあったことは後に丹羽さんがGFPの漏れ出し現象を発見してから気づかれた。ティシュー論文段階からスフィア塊単位で20個から50個に一つのシャーレから三胚葉分化が確認されていた。分化は1000個程度の細胞で構成されている一つのスフィアの中に1個でも多能性細胞が含まれていたら起きうるものです。
この確率の低さについては、これも丹羽さんが再現実験でかなり厳密に計算する実験を行っている。丹羽さんの計算では500,000個の細胞酸浴によって最大30個のスフィア塊ができ、そのうちの2割のスフィア塊が1個から2個のOct4遺伝子を発現しているという結果です。この結果は20個から50個のスフィア塊からやっと一つ三胚葉分化してくるという小保方さんの報告と一致します。
無論、この結果は小保方さんの三胚葉分化実験では確率的に生じ得ることですが、これが若山さんのntES化実験で使われたときにどうなるかということです。ntESというのはまずはクローン胚の中に小保方さんの細胞の核を一個ずつ入れるものです。1000個単位で20も50も行ったものを数セットやるのではない。この膨大なシャーレに関しては小島さんがたまには整理しろと小保方さんに命じた逸話が残されているくらいですね。1000個単位のスフィア塊ですらこれだけの実験を行わないと確認できないものです。
若山さんは当然ティシュー論文の話は知っています。小保方さんの以前の実験ではOct4遺伝子発現細胞はわずかだと知っている。その確率の低さは厳密には知らなくても、仮に1/2であっても小保方さんの細胞核を使う実験なんてできません。20個から50個のスフィア塊に一つなんて言われたら何をクローン胚に入れることになるか分かりません。そんな実験はしませんね。
若山さんのところで光った細胞はそんな数では無かった。西岡さんのガンバレブログに楠本さんがアップされているOoboeさんのパートナー氏が入手された写真です。これもあちこちに保存コピーを確保しておくという意味でここにも貼り付けておきましょう。左側は赤色フィルター画面です。自家蛍光の無いものです。これを最初見せられた時はショックを受けましたね。これだけ光ったら誰でも何物かだと思いますね。
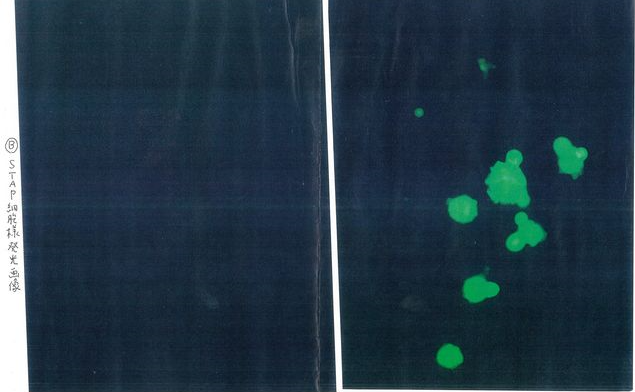
若山さんが酸浴細胞に関してもヴァカンティには何の権利も無いなどと言ったという手記の記事も理解できないではありません。ヴァカンティ研で発見されたのは20個から50個に一つあるかないかの細胞じゃないか。何を見つけていたか分かりはしないぞ。GOFマウスを使わせてやって、赤ちゃんマウスを使えとアドヴァイズしてやって、こんなに「よく光ってるね」と確認できる細胞は理研若山研で発見されたのだと。まあ、アイディアの発端がヴァカンティ研にある位の事は認めてやろうと。ただ、この細胞からはキメラができなかったんですね。
この蛍光スフィア塊の中からいくつか取り出してクローン胚を作る。科学者というのはなんでもやってみるんですね。やってみなければ分からないことを発見しようとしている。推測ではできないからやらないと言ってたら自然界から何か新しい事実を発見するなんてことはできない。思いがけない事実を発見するから発見なんで予期した通りの事なんて、すでに人が知ってるから予期するのであって、発見にはならない。
ここに歴史眼が必要なんですね。自家蛍光かもしれないなんて考えないレヴェルで博士なんてあり得ませんね。実際ここでも赤色フィルター確認されているし、既述している通り、ライヴセルイメージングでも確認している。自家蛍光ではないんですね。自家蛍光でないことを確認しないで実験なんてやるわけがないでしょう。馬鹿かというような話です。そんなことをノフラーというのは喧伝していた。間抜けた話です。あんなヨタ話についてくるマスコミのレヴェル低下というのもひどいものです。チイトハ考えろや。
でも、それは既存の知識で熟慮すればわかるでしょうという話に過ぎない。科学というのは既存でない知識を得ようとするものですから、丹羽さんが後に発見したGOFマウスの細胞の酸浴によるGFPの漏れ出しなどという事実は、この頃の若山さんは知りません。これが歴史眼です。彼がそれを知らない状態で行動しているということを理解しなければならない。
若山さんは小保方さんのOct4遺伝子発現している細胞核をクローン胚に入れたと信じているんです。でも、丹羽検証の結果を知っている我々はそれはただのGFPの漏れ出しで光っているだけで内在性Oct4遺伝子を発現している細胞で無い確率がはるかに高いということを気づける場所にいる。
そしてそこから演繹できる結論は若山さんはただ白血球の酸浴細胞で生き残った細胞の核を使ったクローン胚を作ったに過ぎない確率が高いということです。そして、仮に結果的に若山さんがいろいろ調べた結果が、単なる普通の体細胞ntESであったとしたら、それはただ単に自分の研究で新たな発見をしたに過ぎないということになる。でももし彼が小保方さんの細胞に出会わかったら彼はそんな研究は決しておこなわなかったでしょうね。
彼のntESの胎盤は光ったかもしれません。どうしてかというと、クローン胚はそもそも体細胞がリプログラムしているものです。自然胚と同じく胎盤貢献はもともとするものなんです。ただ、ESとして取り出したときに、自然胚のESとクローン胚のESと同様に常識的には胎盤貢献しないと思われていただけかもしれないんです。ntESの胎盤異常は大変有名な研究課題です。クローン胚というのは確かに成功すると自然胎児と同様に生まれてくるんですが、自然胚には胎盤異常なんてめったにありません。自然胚が胚盤胞になったとき外側のトロフォブラストと内側のインナーセルマスに分かれる。外側が胎盤形成するんです。中に残ったインナーセルマスは胎盤になりません。だからこの段階で取り出したES細胞は胎盤貢献しないんですね。でもクローン胚のインナーセルマスはどうでしょうか。クローン胚には胎盤異常が多発する。そこにはトロフォブラストとインナーセルマスへの機能分離が完全には行われていない可能性も考えられる。するとクローン胚のインナーセルマスを取り出したものであるntESに胎盤への分化能が残る可能性が理論的にはありますよね。
今回小保方細胞を入れたと思い込んでいることから調べてみることになっただけで、結論はひょっとしたら小保方細胞とは無関係にntESの胎盤は光るのかもしれないんですね。そのことは若山さん自身が後に言ってますね。そもそも正直な人なんでしょうね。内心本当の事を知ってもらいたいという気持ちはあるんでしょうね。
尤も笹井さんはああいうことになっちゃったからもはや本当の事すら言えなくなってしまった。あれは笹井さんがいけないですね。自決は無論自己責任です。誰かの所為にすることはできませんが、他者が何も言えなくなってしまいますからね。そういう意味であれは笹井さんがいけないですね。
(如何にしてヴァカンティの手から小保方さんを奪うか)二兎を追うとどこかで破綻しますね。若山さんは自分の研究と、小保方さんに論文を書かせてヴァカンティの手元から小保方さんを奪うという仕事を両方行わなければならなくなった。そもそもはキメラが出来なかったから帰ろうとした小保方さんを2,3か月引き留めて置くだけの軽い嘘だったもので、翌年に小保方さんがはっきり山梨大に行くとか行かないと回答していたらこんな問題にならなかったはずです。
ただ、小保方さんはあの時点で行くという返事はできません。あの時点ではハーヴァード大の所属で給与は正式に米国から小保方さんの口座に振り込まれています。理研ではただの客員で、小保方さんはハーバードから日本へ長期出張してきているだけです。2012年の4月、もしくは2013年の4月から山梨大に行きますとは言えない。ハーヴァードとの約定がどうなっているかは分かりませんが2年契約とか3年契約だと行けませんし、常識的に1年契約の双方異存なきときは自動更新だとしても、ヴァカンティの言によれば論文が出るまでは動けない。小保方さんの願いを入れて日本への長期出張を認めてくれたヴァカンティ先生を裏切ることはできないし、そのバックには常田、大和という恩師の紹介や人脈あっての就職です。でも、行かないという即答はあり得たんですね。
因みに武田邦彦教授は小保方さんを研修生のような理解でおられるが、小保方さんの給料はハーヴァードからちゃんと支払われていますから、理研が支払っているのは客員への謝金だけです。あの部分の認識は間違いです。ただ共同研究協約書のない客員受け入れなので後にそれがいろいろとトラブルの原因になってるんですね。
行かないと断るとどうなるかというと、この共同研究は打ち切りでしょうかね。そもそも若山さんが自分の研究費を使ってハーヴァードの研究を手伝っているという姿そのものがおかしいと誰でも思いますね。現に岸氏はとても怒ったらしいですね。小保方さんが院生のころに若山さんは好意でキメラ実験を引き受けてあげているんですね。小保方さんは当時東京女子医大の院生です。だから理研は研修生受け入れできる。ところが細胞の権利がハーヴァードにあるので、ヴァカンティはその権利を奪われるのを恐れたんです。小島さんを通して、共同研究の形でキメラ実験を行ってくれと頼んだ。研修生なら簡単なんですが、共同研究だなんて、ただキメラ作ってあげるだけの話です。どんな契約文面にするのだということです。若山さんが申し込んだんじゃない。若山さんは断ればよかっただけです。でも日本人の学生で、研修生なら簡単に引き受けられるものを米国が細胞の権利を留保するためにそんなことを言いだしているだけです。若山さんはめんどくさいから、客員受け入れの形で手伝ってやってるんですね。形式的なものです。震災後の受け入れでも若山さんはその延長線上の考え方で小保方さんをひきうけてやっている。ただ、渡米できるようになった時に自分のラボにヘッドハンティングしようとして、結果ヴァカンティが断った。でも小保方さんは若山研のGOFマウスを使って研究したかったから、ヴァカンティに頼み込んで理研で研究できるようにしてもらった。この時の交渉は小島氏と若山さんの話合いと推定されますね。その時に共研究契約書を交わさずに以前通りの単なる客員受け入れにしてしまったんですね。
まだ何も発見はありません。若山さんはヴァカンティが論文が出るまでは自分のラボに所属していてくれと言ったという小保方さんの言を何となく聞いている。小島も同様のことを言ってるんでしょうね。若山さんとしてはキメラを作ってやるだけの話と思っている。共同研究だなんて言われても、自分が望んだことではない。相手が頼んできているんだ。でも理研は日本の会社ですからね。原則理研が望んで共同研究するので無いと認めませんね。若山さんもなんらか理由は考えておかないといけませんよね。共同研究だと研究計画書をださないといけないんですが、我々はそれがあると思っていたら、理研は公開請求に対して無いと答えました。
共同研究の約定が無いことの問題は、GOFのGFPが光り始めた時から顕在化し始めたでしょうね。若山さんはそんな細胞は出来やしないと内心思ってますからね。いつまでこの子を預かってりゃいいんだという気持ちですね。とても中途半端な気持ちですね。どうせできないだろう。できなければヴァカンティとも別れて自分のところにいずれ来てくれるかなとも思うし、どうなるか分からないが、熱心さは買っているんで、自分のところに連れてきたいと思ってますね。現に一度誘ってますからね。これは酸浴細胞とは無関係なんですね。むしろ自分の弟子にして自分の研究を発展させてもらいたいと期待する。
しかし、光り始めた時には何物なのだと思いますね。GFPの漏れ出しなんて考えもしてませんね。Oct4-GFP蛍光=Oct4遺伝子発現という認識です。光ってはいるが、前回も確認してキメラは出来ませんでしたからね。キメラは出来ないだろうなとは予測しているがそれでもいろいろと確認して最終的にできないと結論した。そこからこの細胞をntES化してみようという自分の関心が生まれてくるんですね。岸氏がけしからんと怒るが、そもそもそんな共同研究なんて考えていたわけではない。ただキメラを作ってやるのに便宜的な形にしておいただけだ。まさかあんなに光るなんてわかってたら約定してたかもしれないくらいだ。そんなことはまるで考えてなくて、ただ小保方さんをリクルートしたいなあと思ってただけだ。あくまでも好意で預かってあげている。
ハーヴァードは小保方さんの給与も出張旅費も滞在費も無論全部負担している。でもそれはハーヴァード側の当然の負担で、若山さんが何かを頼んだわけではない。それどころか若山さんは自分の研究費で小保方さんの研究を手伝っている。理研の資材を使わせていることも含めて普通はその負担を契約書で分担して無いとおかしいんですね。単なる客員預かりの届出書で処理されていると理研は言ってるが、社会常識でそんなことはあり得ないことなんですけどね。でも、10年以上保存原則の契約書が保管されていないと言ってますからね。とても不可思議なところです。
常識的には形だけでも契約書は無いとこの客員受け入れは承認されませんよね。因みにこの時小島氏も客員に登録されています。10月には酸浴細胞の実験のための動物実験申請書が副所長だった当時の相沢さん宛に提出されていますよね。あくまでも若山さんの研究として提出されている。若山さんは約定もなく自分の研究として税金から出ている自分の研究予算を使い、日本の税金で運営されている理研の資材を使って、米国の研究を手伝っている。普通は違法行為でしょう。よくこんなことができましたね。契約書は曲がりになりにでも有ると考えた方が理解しやすいんですがね。岸氏が怒ったのはその内容が杜撰だったということだったとしたら理解可能なんですが、最初から何もないとなると、怒って済むような問題に留まりますかねえ。しかも理研の調査では何度もこの研究を共同研究だったと公表している。理研の調査報告書はすべて公文書です。そこに契約書も交わされていないのに共同研究であったなどと書かれていること自体変ですね。
(細胞塊は本当によく光った)
これが事件の深奥に横たわっている真の原因だったかもしれませんね。知られていなかったアーティファクトの存在です。
我々は今AC129の謎を解明しようとしている。こんな時期になぜ若山さんがこんな実験をしたのか、そしてなぜ小保方さんがこの時に渡されたはずの赤ちゃんマウスに関してローザだと書いているのか。そして小保方さんはキメラが作製されたと書いているのに若山さんはなぜキメラを作っていないと答えているのか。更にはなぜ公共データ登録されている試料に129/Svのエピプラスト幹細胞があるのか。
我々はこの頃若山さんが信じ込んでいるよく光っている細胞核のntESであるはずの幹細胞の正体に疑念が出たのではないかとみています。その切っ掛けは西川さんがアドヴァイズしたTCR再構成のPCR検証で自分の予測していた結果が出なかったことではないかと考える。
(西川アドヴァイスの余波)幹細胞は論文通りの作り方であると無論TCR再構成に関してはポリクローナルな集団ですからPCRに掛けると全バンドが出ないといけない。しかし、若山さんはクローン胚の胚盤胞のインナーセルマスを取り出していますから、もともとが一種類のTCR再構成を起こしている一個の細胞が自己増殖しているものですから、そのTCR再構成結果はいろんなシャーレのものを混ぜていない限りは0本か1本か2本のいずれかのバンドが出るので無いといけない。キメラは今は置いておいて、特に幹細胞FLSは全部雄です。これは小保方さんの細胞核を使ってたくさんクローン胚を作って、それぞれを別のシャーレで維持培養しているものを持っていたでしょうが、それらを混ぜてはいないということを意味している。
まず、事実関係の確認です。手記の主張するところでは2,3株にTCR再構成があったという結果だったが、後に調べ直したらどれにもなかったと書かれている。
桂報告書は以下のように書いている。27P。
>>
1)TCR 遺伝子再構成に関する不整合データ隠蔽の疑いについて
(調査結果)
小保方氏は TCR 遺伝子再構成に関する実験を開始し、STAP 細胞を含む細胞塊、一部の STAP 幹細胞に TCR 遺伝子の再構成が見られることを CDB 若山研で最初に報告した。しか し、後に 8 系統の STAP 幹細胞の TCR 遺伝子の再構成を確認したところ、再構成は確認 されなかった。なお、この8系統は小保方氏が継代培養を繰り返していた細胞であった。
さらに、この実験は小保方氏の依頼で、CDB 若山研メンバーによる TCR 遺伝子再構成 の確認実験が行なわれた。しかし、この CDB 若山研メンバーの実験ノートによれば、実 験の結果 TCR 遺伝子の再構成は確認されなかった。
以上のことから、小保方氏は最初の実験でTCR遺伝子再構成があることを報告したが、 後の小保方氏自身の実験、および CDB 若山研のメンバーに確認を依頼した実験では TCR 遺伝子の再構成を認めるに至らなかったことから、実験データに不整合が存在したこと は明らかである。
丹羽氏は 2013 年 1 月に論文作成に加わった際に、小保方氏が継代培養を繰り返して いた 8 系統の STAP 幹細胞の TCR 遺伝子の再構成は確認されなかったと聞いたと説明し ている。さらに、丹羽氏は笹井氏に対して、TCR 遺伝子再構成に関するデータを論文に 含めることについては慎重にすべきとの意見を伝えた。小保方氏の追試が不成功であっ た点に関して、笹井氏らは STAP 幹細胞がヘテロな集団であり、長期的な継代培養により再構成が起っていた細胞が消失したという解釈を採った。なお、Article 論文には、 STAP 細胞を含む細胞塊の TCR 遺伝子再構成については記載されたが、STAP 幹細胞自体 の TCR 遺伝子再構成実験の結果については記載されなかった。
一方、丹羽氏は、Protocol Exchange への投稿は、発表後、この論文ではすぐに再現 性についてクレームがつくと思った。小保方氏のプロトコールでは不十分と考えそれを 詳細にしたものを早急に公表すべきと考えた、と説明した。さらに、当時、小保方氏と 笹井氏はコリジェンダム(corrigendum)で相当に多忙であり、エディターと応答でき る者が必要ということで、自分が執筆した、と説明した。
2014 年 3 月 5 日に Protocol Exchange に公表された詳細なプロトコールの「STAP stem-cell conversion culture」「 2.After 4-7 days of…」のプロトコールの「IMPORTANT」 (iii)に、8 系統の STAP 幹細胞には TCR 遺伝子再構成が認められない、という結果の記 載が存在している。 また、丹羽氏は「若山さんは、最初 STAP 幹細胞の初期のパッセージでは TCR 遺伝子 再構成はあった、と小保方さんから聞いたと言っている」と説明した。
(評価)
TCR 遺伝子再構成に関しては、最初小保方氏が再構成を確認したとされたが、その後 の CDB 若山研メンバー、および小保方氏自身の追試で失敗した。その事実にもかかわら ず、実験結果を自分たちのアイデアに沿うようなものを採用したものの、後に、Protocol Exchange で 8 系統の STAP 幹細胞には TCR 遺伝子再構成が認められないという結果が記 載されたこと、並びに丹羽氏への聞き取り調査における上記の説明から、意図的な隠蔽 ではなく、研究不正とは認められない。 手記の148Pは以下です。
>>
STAP幹細胞は理研の細胞リソース部門に移管し、外部の研究者たちにもSTAP細胞を取り寄せて実験に使ってもらえるようにすることが予定されていた。STAP幹細胞のTCR再構成については、当初若山研のスタッフによって解析が行われた。その時の結果では調べられた8株のうち2株にはTCR再構成があるようだったが、その実験にはコントロール実験がなく、結果の正確さは担保されていなかった。そのため私が後日、自分で確認の実験をコントロール実験と同時に行ったところ、どの細胞からもTCR再構成は観察されなかった。しかし、私が解析したSTAP幹細胞は若山研のスタッフが実験を行った細胞から継代培養の過程で選択がかかりTCR再構成のない細胞だけが生き残ったのかが不明瞭になってしまった。ただ、外部に譲与される予定だったSTAP幹細胞は少なくとも継代培養によって増やされたものであるので、プロトコールにはSTAP幹細胞にはTCR再構成がない、と記載されることになった。桂報告書は相変わらず支離滅裂な報告ですが、
①まず事実関係で手記との間に矛盾がある。手記では最初に幹細胞のTCR再構成確認実験を行ったのはスタッフと書かれているが、桂報告は「小保方氏は TCR 遺伝子再構成に関する実験を開始し、STAP 細胞を含む細胞塊、一部の STAP 幹細胞に TCR 遺伝子の再構成が見られることを CDB 若山研で最初に報告した。」と書き出し、"最初に報告した"と書くことによって、小保方さんが幹細胞の最初の確認実験を行ったかと誤認させるような卑劣なレトリックを使っている。ここはテクニカルスタッフが確認したとちゃんと書くべきところでしょう。小保方さんはその結果を聞いてCD45陽性細胞や酸浴STAP細胞やと合わせて報告したということになる。印象操作をしている。小保方さんが嘘をついているのなら、そのスタッフの実験ノートを証拠として提示しないといけない。
②次に、桂報告書はTCR遺伝子再構成の有無はどういうバンドが出た時に有りで、どういうバンドの出方では無しなのかの説明がない。ひょっとしたら考えてない可能性すら疑われる。少なくとも小保方さんは全バンドが出たときが有りだという理解であることはGel1、Gel2の写真からアーティクルの画像を構成していることから明らかですが、全ラインから同じように出ないとおかしいということに関して、このラボ内での報告の時疑義表明していません。どうして若山さんは幹細胞の確認実験を小保方さんにやらせずにテクニカルスタッフにやらせたのか。これを調査しないといけませんね。小保方さんが自分で最初の幹細胞の実験をしていたらいわばGel3のようなものがあったはずでしょう。
③この時にラボに所属していたテクニカルスタッフは坂出裕子さんと山中香織さんでしたね。前者は太田論文の共著者ですね。今も理研に勤務されている。後者は大阪のエキスポ館でしたっけ。李さんとメールしてたり、ハーイポールなんておっしゃってた方でしたか。コントロールの無いPCR実験を行って、2つにあったと小保方さんに言ったんですね。どうして6つには無かったのか。そんな筈ないでしょ。あったら全部にありますよ。そもそもあったと言ったこと自体どんなバンドがあったのかすら怪しいですね。小保方さんは客員だからラボのプログレスレポート時期に疑念を言い出せなかっただけではないか。この幹細胞は太田ESだと言われているんですからね。最低でも全部にGLバンドが1本出てたはずなんですがね。ESであったらバンド無しはあり得ません。バンド無しはTCR再構成があるということです。ただし、その場合検体はモノクローナルな集団だということを証明する。
④桂報告は「なお、この8系統は小保方氏が継代培養を繰り返していた細胞であった。(段落) さらに、この実験は小保方氏の依頼で、CDB 若山研メンバーによる TCR 遺伝子再構成 の確認実験が行なわれた。しかし、この CDB 若山研メンバーの実験ノートによれば、実 験の結果 TCR 遺伝子の再構成は確認されなかった。 」と書くが、手記では小保方さんは継代培養を繰り返していた細胞の再確認は依頼してない。「さらに、この実験」と言ってるのは最初の実験なのか。最初の実験を小保方さんがスタッフに依頼してたのだったら、最初の実験はスタッフだと知ってることになるではないか。どうしてまずそれを書かないのだ。それとも確認実験を小保方さんがスタッフに頼んだというのか。それなら小保方さんが手記に書いていることは嘘になる。手記には自分でコントロールを取ってやり直したと書かれている。
⑤「この CDB 若山研メンバーの実験ノートによれば、実 験の結果 TCR 遺伝子の再構成は確認されなかった。 」と書かれている実験ノートは公開されなければならない。なぜなら、この実験が最初の実験であるならば、このラボメンバーは小保方さんがラボで幹細胞にもTCR再構成があったと発表した時に嘘だというでしょう。同じ場所にいるのにそんな確認もできないか。仮に小保方さんが手記に書いている再確認時に誰かに手伝ってもらったものの事だったらTCR再構成は無かったと小保方さんが書いていることと何も矛盾が無い。これを手伝ったのは寺下さんでしょう。手記に書かれている最初の実験時のスタッフではない。
⑥「(評価) TCR 遺伝子再構成に関しては、最初小保方氏が再構成を確認したとされたが、その後 の CDB 若山研メンバー、および小保方氏自身の追試で失敗した。」という認識は最初の実験を小保方さんが行ったと誤解していることが明確な書きようである。間違ってるんです。最初、若山さんは幹細胞のTCR確認を小保方さんにやらせなかったのだ。手記はスタッフと書いている。
(若山さんの行ったTCR再構成確認PCR実験)幹細胞のTCR再構成確認実験は若山さんとテクニカルスタッフの間で行われた。この時の確認がFLS8株であるはずはありません。若山さんは成功したいくつかのntESラインの維持培養シャーレを持っている。CD45陽性細胞というのは白血球の共通抗原ですから、いろんな種類の白血球が混在している。好中球・好酸球・好塩基球・リンパ球・単球とあって、リンパ球にもT細とB細胞があって、無論TCR再構成のあるのはT細胞だけです。若山さんにしても自分のntESのそれぞれが何であるかは確認しておきたいでしょう。西川さんのアドヴァイスは若山さんの研究にも影響を与えたんですね。
でも、TCR再構成確認では若山さんのntESの由来細胞が何であるかを決定することはできませんね。西川さんが小保方さんにアドヴァイズしたのはポリクローナルな細胞集団であるはずのSTAP幹細胞集団ならTCR再構成の全バンドが出るから、STAP細胞がT細胞を含んでいるリンパ球、もしくはもっと広い範囲で白血球由来であることは証明可能だというものです。T細胞が少しでも含まれていたら他の細胞由来のものもあったとしても、TCR再構成の全バンドが出るから、少なくともT細胞もSTAP幹細胞になっていることは証明できるのだということです。PCRは目的の遺伝子断片を増幅しますから少しでもあれば全部拾ってくる。でも最初から存在しないゼロはどれだけ増幅してもゼロです。
若山さんは幹細胞をntESとして作っていますから、全バンドが出ることはないということを知っているはずです。はずだというのは、専門が離れすぎていていて、西川さんなどに比べていつも考えていることではないということから、STAP実験の各種試料のTCR再構成のバンドがどういう具合に出るものなのかということに関して、深く考えたかどうかが分からないということです。小保方さんも若山さんも医学系ではありません。常識的なことは当然学んではいるが、常時免疫のことを考え続けている仕事ではありませんね。
我々ど素人がちょっと調べただけでも、小保方さんのTCR再構成の概念図はミスリーディングなものです。笹井さんも丹羽さんも概念図だからいいだろうと判断したかもしれませんね。J2の7つのセグメントのうちの6番目は偽遺伝子なんですが、それも含めて1から順番に番号が打たれている。そして彼女は6番目のところでプライマー設定していると図示している。つまり7番目は後ろに残されていると。ところが、アルイミオウジ氏が紹介している論文から、小保方さんの使っているプライマーは有名な河本論文のプライマーと同じであることが判明した。河本論文では偽遺伝子を数えないでJ6の後ろで切り取っているのです。これですと出てくるべきバンド数が変わってきますね。ただし、バンドの数は又、遺伝子の距離が近いと重なってしまうということもあります。なかなか単純でないんですね。特にこのPCRに関しても専門家がいて、彼らに頼むと厳密にバンドが出て来るようなテクニックがあるようですね。若山ラボで行われたPCR実験は手技の稚拙さも考えなければならないようなんです。つまり、実験はちゃんとした技術で行われているかという問題まであるということです。
因みにアルイミオウジ氏は我々のしたらば掲示板の書き込みを荒らした者と同様に我々のブラックリストに上がっているスピン書き込み者です。いずれ11jigen や世界展望なんかと同様に、その素性に関してこの便所の落書き新聞で検討することになるでしょう。スピン屋には3種ある。若山ラボ関係者、関与マスコミ関係者、文科省関係者、厳密には変質者を入れて4種かもしれませんね。
ともあれ、このTCR再構成確認は小保方さんを含めて若山ラボには難しかったようです。加えて、若山さんは論文を通したくないという姿勢で小保方さんに接していますからね。実験は厳密でないほどいいのだという裏事情がある。論文がこの証明で通っては困るんです。小保方さんは2つにはTCR再構成があったと手記に書いている。これって幾分かでも専門的に考えることのできる人だったらおかしいと思います。どうして8ライン全部に出ないのか。小保方さんは気づいていません。そもそもどんなバンドが出ていたのかも誰も書いてない。桂報告さえ出るべきバンドについて言及していない。
唯一分かっていたのは石井さんで、あの図は白線さえ入れていれば何も問題なく、かつ、ちゃんとバンドが出ていると言っていますね。分かっているんです。ここに2つにだけある幹細胞のPCR結果があったら、石井さんは変だということを指摘したでしょうね。それは白線を入れるというような些末なルールの問題ではない。この幹細胞と呼ばれているものは何だと指摘することになったでしょう。それはキメラのgelを見ても言わねばならなかったはずのことです。変だと問題提起しないといけなかったでしょうね。ただ、これは丹羽さんと笹井さんが変だと気付いて何かの手違いだと考えたので、論文から外させている。だから問題にできなかったんですね。
話をAC129に戻しますが、AC129という幹細胞は試料として残されているだけの話で、この頃若山さんが何のために129/Svを使って小保方さんにSTAP酸浴塊を作らせているのかは若山さんのストーリーに従って考えることはできません。彼は既にたくさんの嘘をつき不審な言動をしていることが判明しています。我々が彼の説明を真に受けて考える理由などありません。小保方さんはアーティクルに 129/Sv carrying Rosa26-gfp (2 of 2) と書いています。そして論文は共著者は全員読んでいる。
>>
STAP stem-cell conversion culture
For establishment of STAP stem-cell lines, STAP cell clusters were transferred to ACTH-containing medium[36] on MEF feeder cells (several clusters, up to a dozen clusters, per well of 96-well plates). Four to seven days later, the cells were subjected to the first passage using a conventional trypsin method, and suspended cells were plated in ES maintain medium containing 20% FBS. Subsequent passaging was performed at a split ratio of 1:10 every second day before they reached subconfluency. We tested the following three different genetic backgrounds of mice for STAP stem-cell establishment from STAP cell clusters, and observed reproducible data of establishment: C57BL/6 carrying Oct4-gfp (29 of 29), 129/Sv carrying Rosa26-gfp (2 of 2) and 129/Sv × C57BL/6 carrying cag-gfp (12 of 16). STAP stem cells with all these genetic backgrounds showed chimaera-forming activity.STAP幹細胞は小保方さんの作った酸浴STAP細胞をACTH-containing mediumに4~7日オンフィーダーで培養した後、通常のES維持培地で継代するもので、このプロトコルは無論若山さんが小保方さんに伝えたもので、小保方さんはこのプロトコルでできていません。そのことを論文リヴァイズ中に若山さんに相談したら、人が変わるとできないこともあると言って、やって見せようともしなかった。小保方さんはそれを丹羽さんに相談したら若山さんが絶対の自信を持っているんだから信じるべきだと言ったと手記に書かれていて、手記の記載が嘘なら丹羽さんが指摘するでしょうからね。丹羽さんも若山さんを信じていて、後に多忙だった小保方さんの代わりに口伝されたとおりのプロトコルを書いあげている。信じていたからこそ、若山さんが取り下げを主張した時に「梯子をはずされたんや」と口をついて出たんですね。このとき丹羽さんは直ぐに山梨に出かけて行って若山さんに会っています。後の丹羽さんの記者会見で、この件について、若山さんはESを入れられたと信じ込んでいるもんでと答えましたね。この時に若山さんはESだったと態度を翻したんです。
因みに笹井さんと丹羽さんはヴァカンティの米国特許仮申請の期限が迫っていることがあって、小保方さんに信じるべきだと言ってるんで、確認すべきことだということは分かっているんです。だからこそ丹羽さんは急ぐ中でもキメラと幹細胞のTCR再構成確認実験の記述は外させた。これだけでも科学者として大したものだと思わないとけないでしょうね。他のことは勤め人として業務命令に従って行っていることですからね。科学に門外漢のど素人でも理解可能なことですね。組織というのは命令一下各人が歯車になり切って行動しないと仕事になりません。各人が社長のように思い思いの判断で行動したら組織ではなくなる。当たり前のことですよね。
(「舐めてますね」。これ。)アーティクル論文には書かれていませんが、丹羽さんはオンフィーダー培養時点で使われた細胞はキメラと同じくナイフ切り分け塊だということを後のプロトコルに書いています。
>>
The STAP cell cluster was isolated, dissected into small pieces as in the case of injection into blastocysts as shown in Figure 4a (Obokata et al. Nature, 2014a), and seeded on mouse embryonic fibroblast feeder cells in the ACTH medium.論文のプロトコルには無い情報です。若山さんに確認して追加しているのではありませんか。丹羽さんと若山さんは知人ですね。世界からの情報で再現が取れないので、プロトコルを書くということになって、若山さんに手技を聞いたんでしょうね。その言い訳の中に幹細胞もキメラと同様にナイフ切り分けで行うのだと答えたんでしょう。重大な情報ですね。小保方さんはマニピュレーターを使えませんから自分の細胞塊をあのようにカットはできないんです。小保方さんにできるのはESの培養と同じようにトリプシンでばらして培養するか、カットせずにスフィア塊ごと培養するかです。彼女がどうやったのかは分かっていませんが、カットしてないことだけは分かります。もっともカットしてできるのなら塊のままでもできますね。これはキメラではありませんから大きすぎるなどという問題も無い。カットするのが手技だというのも妙な話です。
そもそもこんなことは小保方さんが聞いているんだから教えればいいだけの話ですね。若山さんが教えない理由がありませんね。仲間ではないのか。ヴァカンティに知られたくないのでしょうか。そういうことですと、一体どうして共著者になんかなっているのだという話になってしまいますねえ。それも責任著者で通常直接の先生がなるものだ。キンガの論文の責任著者は当然ムーさんです。若山さんとヴァカンティはライバル同士で共著者になってるのか。
このプロトコル発表後すぐに遠藤氏による「舐めてますね」という書き込みがあった。同じ3月5日です。お前はいきなりどこから来ているんだというような書き込みです。何が舐めてるんだ。お前は何者なんだということですね。その後すぐに若山さんが取り下げだと騒ぎだした。3月10日です。
お前たちの関係を言って見ろ。東北大繋がりなのかな。
>>
2014/01/28 STAP論文記者会見発表
2014/01/29 岡部氏スペルムエッグ書き込み
2014/01/30 西川さんブログ書き込み。駅に記者が張っているからタクシーで来るよう理研から電話。
2014/02/02 若山氏クムリナ書き込み
2014/02/04 論文発表から1週間がたったころ日本で一番大きな生物学分野の学会から竹市所長に連名メール(142P)。ティシュー誌にゲルの使いまわし疑惑の指摘。確認中画像の間違いを発見。ゲルの方は小島氏がハーバード側の責任と謝罪。
2014/02/13 平成26年2月13日、独立行政法人理化学研究所(以下、「研究所」という。) の職員らの研究論文に疑義があるとの連絡を受けた研究所の職員から、役員を通じ て監査・コンプライアンス室に相談があった。監査・コンプライアンス室長は、「科学研究上の不正行為の防止等に関する規程(平成24年9月13日規程第61 号)」(以 下、「規程」という。)(参考資料)第 10 条第 3 項に基づき、当該相談を通報に準じて取扱うこととし、規程第 11 条に基づき、同日より同年2月17日の間、予備調査を実施した。平成26年2月20日から同年3月31日までの間、関係資料の収集・精査及び 関係者のヒアリングを行った。
2014/02/14 11次元の博論画像流用指摘
2014/02/?? ネイチャー・インタビューにて若山氏「STAP細胞は小保方さんに教えてもらってできた」(210P)
2014/03/?? NHK藤原淳登記者から携帯に電話が頻繁に来る。
2014/03/05 丹羽氏プロトコルイクスチェンジ報告。kahoの日記「なめてますね、これ.」。
2014/03/09 小保方さん共著者ともネイチャーに訂正を提出した。
2014/03/10 竹市氏論文撤回を強く勧める。林GDから撤回しろという強い文面のメール、丹羽氏山梨に行く。山梨大若山氏「STAP存在確証なし」(152P)
2014/03/11 東北大大隅声明
ナイフ切り分けしてその中にESが入っていたのに気づかなかったと。そしてESなら簡単に増殖したであろうに、小保方さんができないと訴えているにも関わらず、人が変わるとできないなんて、あなた責任著者じゃないのか。論文を通したくない人なのかね。
論文に作られたと記されているSTAP幹細胞は以下の3種です。
①C57BL/6 carrying Oct4-gfp (29 of 29)
②129/Sv carrying Rosa26-gfp (2 of 2)
③129/Sv × C57BL/6 carrying cag-gfp (12 of 16)
①は常識的にはGLSですね。③はFLBもしくはFLSです。若山さんの持ち出しリストを再掲しましょう。数があってないですね。
そもそもキメラはともかくとして、STAP幹細胞が若山さんの口伝のプロトコルでできないというのは変なんですね。隠している手技があるから他人にできないんです。論文の筆頭著者ができないままに論文を書かせたのは若山さんの責任ですね。論文の共著者は論文が通るように努力するのが当たり前です。小保方さんが太田ESを渡していたら簡単にできたでしょうよ。若山さんも簡単にできたはずだ。手技なんてどこにありうるのでしょうかね。STAP細胞を持って山梨に来い。プロトコル通りに簡単にできるぞと言わないと変でしょ。ほら、できるじゃないか、安心して提出しろと言わないとおかしいでしょ。ESを渡されていたのだったら簡単にできたはずだ。キメラほどの困難もない。隠すような手技なんてありえないじゃないか。それでも騙されていて自分では手技があるのだと信じていたんだというのなら、cumulina書き込みの手技と論文プロトコルの違いを説明しなさいよ。
実験でできてるのは若山さんだけじゃないか。彼が作って見せなくて誰にできるのでしょうかね。そうしなかったのは、小保方さんが細胞を持ってきてもできないと分かっているからだ。一旦クローン胚に入れないとできませんよね。目の前ではやってみせられないんですね。
(ローザという言葉が出てくる理由)AC129は若山さんの持ち出しリストには2つ作って2株樹立と書かれている。でも他の試料もすでに数がおかしいというのが判明していますから、これも真に受けられませんね。竹市さんが「細胞の出所が分からなくては特定の結論を導き出すことはできない」と2014/6/5の埼玉和光市での会議にテレビ会議出席した時の言葉の通りの結末ですね。(『STAP細胞 事件の真相』79P)
[②129/Sv carrying Rosa26-gfp (2 of 2) ]は小保方さんが論文に書き込んでいる幹細胞で、キメラ確認もされていると書かれている。これが若山さんの持ち出しリストのAC129であるという証拠は何もない。更に、木星リストに並んでいるAC129の由来も又明確になっていない。AC129の入っていたボックスがリスト以外となっていることに注意が必要です。

この細胞は丹羽さんが持ち出しています。

2014/6/2に解析を受けたとされているAC129-1,2がこれです。(『STAP細胞 事件の真相』80P)
この件に関しては既にOoboeさんのパートナー氏が検察に調査依頼されていますね。
この問題は旧DORAブログでも扱われています。彼は今この問題には関心を失ってるようです。ブログの引っ越しで失われる可能性もありますから、資料保存のためにもここにデータ保管しておきましょう。
>>
A.小保方冷凍庫保全リストに「AC129-1」はない。
2016/5/16(月) 午前 10:27 日記 練習用
ちょっと気づいたんだが、小保方冷凍庫保全リストに「AC129-1」に該当するものはない。桂調査委の報告書には、「小保方研フリーザーに保管されていたSTAP幹細胞AC129-1について解析を行った」と書かれているのだが。これについて、理研広報に尋ねたところ、返事は「わからない」だった。まあ、電話口では即答できないということだろう。あらためてメールで質問を送ってくれというから、そうしたが、「質問によっては答えられない場合もある」という。まあ、それならそれで仕方がないが。疑惑は深まると思うけれども。
B.木星さんへ‥‥もう一度、リストの件。
2016/6/16(木) 午後 0:22 日記 練習用
もう一度、リストの件についてですが、私の新たに入手したリストには、次の4種類が載っていました。
(1)日付のないリスト。
(2)4/14日にリスト化されたもの。
(3)5/14日にリスト化されたもの。
(4)7/19日にリスト化されたもの。
この四つのリストには、それぞれ全く別々の試料が記載されています。合わせると膨大な量です。全部で190項目ほどになります。
AC129-1 AC129-2は、このうち(1)に記載されています。
しかし、それ以外にも、見どころ満載のリストです。この四つのリストは絶対に必須です。必ず入手してUPしてください。もちろん、すでに申請中だと思いますが。「小保方研冷凍庫、保全した試料全ての帰属がついた後の全リスト」‥‥と情報公開窓口に言えば、通じると思いますけど。木星さんがリストを集めている頃の話ですが、この時点ではMTAの問題などまだ解明の途に就いたばかりの時期で、五里霧中です。今となっては事後MTAの約束であったろうことは調べがついていますが、この事後締結の打ち合わせ中に双方でのサンプルのやり取りが疑われるんですね。この件に関しては結局今に至るまで調べはついていません。このサンプルの管理責任者は片山さんなんですが、彼の関与していないところでリスト外からもたらされているサンプルがあって、AC129はそこに入っていたんですね。
我々は若山さんはサンプルの中身をあれこれと入れ替えてると推定しています。特に先に挙げたGOFESの遺伝子異常は若山さんが入れ替えてなかったら小保方さんが犯人になるというくらい決定的なものです。小保方さんが犯人で無いということが証明されると、逆に細胞の中身の入れ替えが確定してしまうような証拠なんですね。あの遺伝子異常はGOFマウス自体には無い異常だとされているので、通常培養の体細胞分裂でたまたまできた異常細胞がその近辺で自己増殖した部分をたまたま分割して継代して行ったものでしょうね。
我々はあのGOF-ESは中身が若山さんによってGLSに入れ替えられていると推定していて、GLSはソート後のGLを培養しただけのもので、GLは小保方さんの作ったGOFマウスの酸浴蛍光細胞のクローン胚から誘導したntES細胞だと考えている。
GLを継代していたのは若山さんですから11/25辺りから2/2辺り過ぎまでほぼ2か月間以上植え継いでいる時に遺伝子異常の塊だけが偶然残されたということですね。GLS1~13の全てに異常があった。継代のやり方で全部に同じ異常が入ったのか、元々のntES化によって最初の段階て発生していたものなのか、分かりません。継代株分け中に13株全体に分散してくというのも考えにくくはありますがね。それよりも酸浴で異常の入っていた細胞核からのntESだとシャーレ全体が同じ異常で占められるということが起きやすいでしょうね。
そしてこの理屈は学生のもntESだったんですから若山さんが犯人でも小保方さんが犯人でも同じことが起きる。では太田ESを小保方さんが入れてないと分かった以上、こちらも犯人は若山さんだ。するとサンプルは必ず入れ替えられているということになるんですね。
因みに培養変異と新たなSNPs獲得を理論的に混同している人々がいますが、SNPsはジャームラインに乗ってきたものを言うんです。しかも例えば129/SvJというマウス系統のSNPsというのは何万年も蓄積してきていて、この近交系マウスをジャクソン研究所が近交系マウスとして確立した時のSNPsを言うんです。これが世界中で使われて、その後何十年経過したところで新たに加わるのはわずかで、遺伝子解析でSNPs分布を調べるときに問題になるほどの数ではありません。培養変異というのは体細胞分裂です。ここで発生している異常は体内で発生している癌細胞なんかと同じで原則遺伝はしない。遺伝するのは生殖細胞での異常だけです。キメラを作ってその異常細胞が生殖細胞に入ってかつ生殖能力を持っていると遺伝形質となる。参考までに注しておくと、キンガの実験ではムーさんの細胞はキメラにしても生殖能力はありませんでした。若山さんのntESは既に畜産応用されていて、精子や卵を採るために4Nキメラを作ってジャームラインを作ります。
後にAC129とされたSTAP幹細胞と小保方さんが論文に書いている129/Sv carrying Rosa26-gfp (2 of 2)とは当初小保方さんに説明されたものとは違う目的の実験ではなかったかと考えて見るわけです。そう考えると、若山さんはなぜRosa26-gfpを使って実験しようとしたのかが問題として浮上してくる。このマウスはCre-LoxPシステムと組み合わせて特定部位でGFP蛍光させるマウスで、一般的に何の遺伝子情報も無いと推定されているRosa26と呼ばれている遺伝子座にGFPを挿入してあるところからその名を負っているマウスです。
丹羽さんが検証実験で使ったのがこのマウスでした。STAP細胞の由来確認のために使おうとした。本当は小保方さんがSTAP論文で使ったのは主に白血球で、かつFACSでT細胞選別までしていますね。でも小保方さんはハーヴァード以来三胚葉由来組織の全てからスフィア細胞を採取して試験管内で三胚葉分化させています。血液以外の体細胞を材料にして若山さんはキメラや幹細胞を作ってはいないはずですが、彼女は自分研究は連続しているものだと考えていますから、物理刺激でもできると書いているんですね。
>>
STAP by exposure to other external stimuli
To give a mechanical stress to mature cells, a pasture pipette was heated and then stretched to create thin capillaries with the lumens approximately 50 μm in diameter, and then broken into appropriate lengths. Mature somatic cells were then repeatedly triturated through these pipettes for 20 min, and then cultured for 7 days. To provide a heat shock, cells were heated at 42 °C for 20 min and cultured for 7 days. A nutrition-deprivation stress was provided to mature cells, by culturing the cells in basal culture medium for 3 weeks. High Ca2+ concentration stress was provided to mature cells by culturing cells in medium containing 2 mM CaCl2 for 7 days. To give a strong stress by creating pores in cell membranes, cells were treated with 230 ng ml−1 streptolysine O (SLO) (S5265, Sigma) for 2 h, then cultured for 7 days. After each treatment, the ratio of Oct4-GFP-positive cells was analysed by FACS.そして丹羽さんは血球を使わずに脾臓、心臓、肝臓を使って追試の実験をしたんです。リンパ球は小保方さん本人が担当しましたね。丹羽さんはこの時にアルブミンクレを使いました。
(そもそもどうして細胞追跡を行わねばならないのか)キメラ胚に入れたのはCAG-GFPマウスの酸浴リンパ球です。キメラ胚自体はICRでGFPのないものです。2Nであれ、4Nであれ、キメラが光ったらドナー胚がキメラ形成したのだということは明らかではありませんか。どうしてTCR再構成だの、ローザなんてことが関係するのか。それは最初の論文査読に原因があって、ESの事故コンタミか、はっきりとは言わないが捏造だろうと疑われたからですよね。こんなの論文としてずっとどこかににぶら下げていたらいつか本当なら誰かが追試するので、発見事実は動きません。無理にネイチャーに信じてもらわなくてもいいですよね。ただどこかの論文には掲載して証拠は残しておかないといけない。
ヴァカンティは本当だと信じていますから、これが本当なら三大誌に載るはずだと考えて自分の主催のティシュー誌に載せようとしなかった。これもまた当然ですよね。
それで小保方さんが仕方なく若山さんと相談の結果西川さんのアドヴァイスを受けに行った。ESではないということを証明しないといけないわけです。ドナーと同じ系統のマウスのES細胞があったら捏造は可能です。だからそんなものは無かったと書くと同時に、口で言って信用されるくらいなら、GFPが光っていることだけでトレース証明は十分のはずなのですから、それではいけないわけです。ESでは無いと証明しないといけない。だからESではなく、リンパ球だということを証明しようとしたわけです。ESは卵ですからまだ未分化な細胞で血液になんかなっていませんからTCR再構成なんてありません。小保方さんのプライマーで挟んだらGLバンド1本しかでません。キメラになっていたら数本のバンドが出るはずだ。或いはSTAP肝細胞なら全バンドが出る。
でも、ここに論理の落とし穴もありましたよね。キメラの場合入れたドナー細胞がどの組織に行くかは分かりません。又何個の細胞が一つの組織に入り込むのかも明確ではありません。厳密な細胞追跡法ではありませんよね。ただ、バンドが出さえすれば少なくともT細胞がキメラになっているという証明にはなる。ただ、この証明はいろいろと難しいですね。GFPが光っているものがTCR再構成を受けているという二つを同時に証明しないといけない。幹細胞では簡単ですが、キメラでは難しい。
それに対して丹羽さんが行ったのはそもそも一からやり直すのですからリンパ球でなくていい。西川さんに当時与えられていた条件は外して構わないわけです。リンパ球ではありませんからTCR再構成は使わなくていい。でも、GFPが光りかつESではないという条件はクリアしないといけない。
実際に使われたのはアルブミンクレマウスとローザマウスのF1です。このマウスは肝臓だけが光っている。卵は光りませんからそのES細胞を作ってキメラにすると肝臓だけしか光りません。丹羽さんはこのマウスの肝臓を酸浴させたものをドナーにしたんです。これをキメラにしたら全身に光るドナー細胞が分布しますね。これでESでないことが証明される。ただし、このキメラは出来ませんでしたね。
若山さんは論文は通したくないんです。丹羽さんがやろうとしたような目的でローザマウスを使ったとは思えません。別のことをやろうとしていたはずです。自分の実験目的のはずです。彼はなぜローザマウスを使おうとしたのか。或いは使っているかのごとくに小保方さんに言ったのか。
事実だけをまず確認すれば彼は129/SvのSTAP細胞を小保方さんに作らせたということです。彼がそれを必要としていた。後のストーリーですと、マウス背景がSTAP形成に与える影響を調べる実験だったということになっている。でも129/SvであったはずなのにAC129なる幹細胞は「僕のマウス」ESでしたね。しかも最後には若山さんしか持っていないF1ESの1だった。129/SvのES細胞は若山研にあるということにはなっていません。今までESを使い分けて来た小保方さんはとうとう何の関係も無い「僕のマウス」ESを若山さんに渡して、幹細胞ができるように捏造したのだと。馬鹿か。
マウス背景がSTAP形成に与える影響を調べる実験だというのに、当該ESがなかったからって小保方さんが系統の違うESを使ってでもできたことにする捏造をしたのだと。
これって、できなかったら、129/Svだとできないのだということになって困るような実験なんですか。背景を違えて捏造しなければならないような実験ですか。こんな馬鹿な捏造者を想定する頭って、どれほど悪い頭なんでしょうかね。きっと博士号を後から取り消される以前に博士課程には進めない頭ではないんですか。そもそも大学入試以前の頭かな。
- 2019/09/10(火) 11:30:30|
- AC129
-
-
| コメント:0
(画像の取り違え)この2Nキメラ実験と同時に4Nキメラ実験も行われている。Article Extended Data Figure 7-d図です。
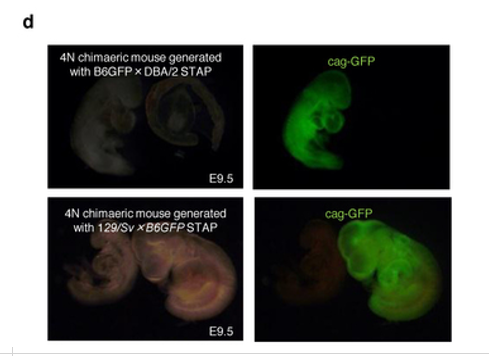
そのリジェンドです。
>>
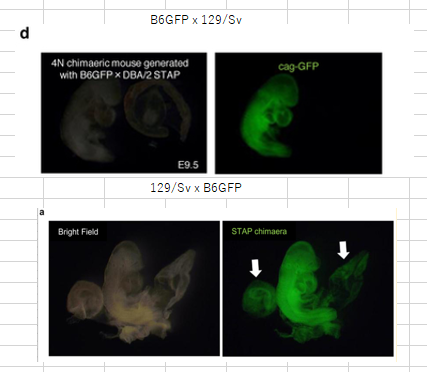
d, 4N embryos at E9.5 generated with STAP cells derived from F1 GFP mice (B6GFP and DBA/2 or 129/Sv). B6GFP, C57BL/6 mouse carrying cag-gfp.Article Extended Data Figure 7-bにある2N実験と対応したマウス背景の4Nです。雌雄をかき分けていることに注意が必要です。それからこれは、2012年2月の実験です。「若山氏の実験ノートでは、この キメラの作製は 2012 年 1 月終りから 2 月はじめにかけて行なわれていた。(桂報告書10P)」のでした。
ところが、この4Nキメラがレター論文の胎盤蛍光写真として使われていると、またもや若山さんが妙なことを言いだした。そのLetter Extended Data Figure 7-aが以下の写真です。
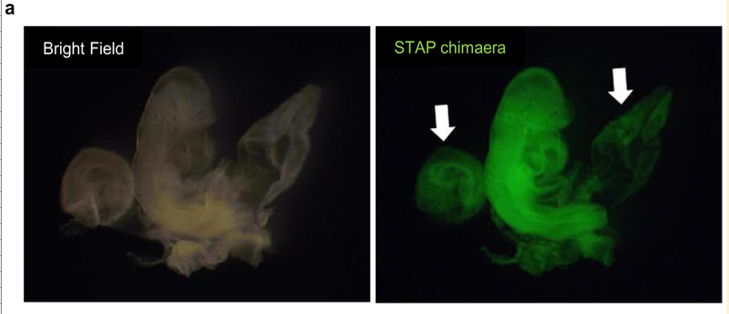
リジェンドは以下です。
>>
a, Chimaeric mouse with STAP cells derived from CD45+ cells of B6GFP × 129/Sv mice (B6GFP, C57BL/6 line with cag-gfp transgene). Arrows indicate a placenta and a yolk sac. これは胎盤が光っているから免染で確認してくれと2012年の3、4月頃に若山さんが小保方さんに指示した時のキメラ胎児胎盤のはずなんですが、それを2月に撮影されているはずの上の4Nキメラの写真と同じものの別角度の写真だと言い出した。手記の99Pです。
>>
若山先生が、スフェア細胞からのキメラ胎児だけではなく胎盤も形成しているようだと報告された。ES細胞はキメラマウスを作製しても胎児の組織を形成することはできるが、胎盤の組織は形成することができないため、若山先生の発見はスフェア細胞がES細胞のの多能性を超える分化能を有していることを示唆していた。スフェアからのキメラマウスの胎盤だというものをいくつか渡され、「(スフェア由来の細胞が胎盤に存在しているかを証明するために)組織学的に解析して欲しい」と指示を受けた、。後日、解析した胎盤内にはGFP陽性細胞が存在していたが、若山先生は私に依頼した解析結果を待つことなく、ご自身の発見から着想を得たようで、2012年4月頃には、TS細胞(Trophoblast Stem Cells:栄養膜幹細胞)と呼ばれる、胎盤を形成する能力のある幹細胞株を樹立する培地でスフェアを培養すれば、TS細胞様の幹細胞株も樹立できるのではないかと、すでに実験を開始されていたようだった。小保方さんの証言では若山さんが胎盤が光ると言い出したのは2012年の4月以前頃ということになる。このためのコントロールESである129B6F1 ES1は2012/4/19に培養開始されている。それ以前に胎盤を小保方さんに渡している。STAP細胞を作り10日前後のキメラを帝王切開して取り出すので20日くらいは前から始まっている実験で3月の末から4月の始めなのである。少なくとも2月の実験分ではない。2月のキメラは2月中に帝王切開それている日程になる。小保方さんはまだ最初のネイチャー論文さえ提出していない。2012年2月頃の写真と胎盤が光ると言い出して小保方さんに胎盤を渡した3、4月頃の写真が同じだと言う。胎児胎盤は生ものです。
若山さんの書いたネイチャーへの取り下げ理由書です。
>>
(2) Extended Data Fig. 7d in the Article and Extended Data Fig. 1a in the Letter are different images of the same embryo and not, as indicated in the legends, a diploid chimaera embryo and tetraploid chimaera embryo.この英文は意味が分かりません。 not, as indicated in the legends, a diploid chimaera embryo and tetraploid chimaera embryo.ってどう解釈したらいいのか。桂報告書の同一の件を見てみましょう。21Pです。
>>
4)Letter Extended Data Fig.1a について
・2N キメラの写真ではなく、Article Extended Data Fig.7d と同じ 4N キメラ胎児胚 の写真の疑いがある点(論文撤回理由 2)(これについては、2014 年 5 月 10 日に著者か ら報告、5 月 21 日に報道されている)
・この写真で胚の一部を胎盤と誤同定している可能性がある点 (Letter Extended Data Fig.1a が2N キメラの写真ではなく、Article Extended Data Fig.7d と同じ 4N キメラ胎児胚 の写真の疑いがある)と主張しているようなんですが、そもそもLetter Extended Data Fig.1a が2N キメラであるなどという記載は論文には無い。若山さんの英文をそう解釈することも又困難で、何を言ってるのか分からない。加えて桂報告書が更に妙なことを言いだす。
>>
(調査結果)
4N キメラ胚であることは、マウス胚撮影に用いた PC に残存する写真(2011 年 11 月 28 日撮影)と若山氏の実験ノートから確認できた。論文の図の説明には 2 つの矢印があっ て、胎盤と卵黄嚢とされているが、専門家の意見によれば 2 つとも卵黄嚢である可能性 が高い。おいおい、2011年11月28日って最初のキメラ成功実験の写真じゃないか。この最初の成功キメラは保存されてない。同時にできたとされていたF1の幹細胞もない。ところがここに突然写真が出てきた。それだけでも驚きなのに、最初の成功時に胎盤が光るなんて話は出てないはずなのに、2011年の11月の写真と、2012年2月の写真と、2012年3、4月の写真が全部同じだという。タイガイニセエヤ。
因みに若山さんの英文をグーグル翻訳機に掛けてみると以下のようになる。
>>
記事の拡張データ図7dおよびレターの拡張データ図1aは、同じ胚の異なる画像であり、伝説に示されているように、二倍体キメラ胚および四倍体キメラ胚ではありません。
Articleが記事、legendが伝説になっているが、そこは機械翻訳なんだから論文と説明に直せばいいだけなんですが、構文的に直訳すると必ずこうなります。つまり、二倍体キメラでも四倍体キメラでもないと。でもキメラって常識的にはどちらかしかありません。学者とか教師というのは職業上の習性があって、本当のことは言わないまでも、嘘がつけないということがあるんですね。事実を曲げることができない。この文意を通そうと思ったら、この胚はキメラではなくクローンだという解釈もできるんです。ただ、証拠がないからね。
桂報告書はこう言うんです。
>>
(調査結果)
4N キメラ胚であることは、マウス胚撮影に用いた PC に残存する写真(2011 年 11 月 28 日撮影)と若山氏の実験ノートから確認できた。論文の図の説明には 2 つの矢印があっ て、胎盤と卵黄嚢とされているが、専門家の意見によれば 2 つとも卵黄嚢である可能性 が高い。
(評価)
2N キメラか 4N キメラかは、論文の重要な論点とは考えられず、過失による可能性が高 いと判断した。STAP 細胞の胎盤への寄与は、Letter の論点として重要であり、研究の価 値を高めるために強引に胎盤と断定した可能性があるが、調査により得られた証拠に基 づき認定する限り、研究不正とは認められない。なお、図の説明にある「B6GFP×129/Sv」 は、最初にメス、その後でオスの遺伝的背景を書く通常の表記法では「129/Sv×B6GFP」 が正しいが、不注意による間違いと思われる。 若山さんは正直な人であくどい嘘をつけない人のようです。彼は助手の採用条件を言えるまでの期間ちょっとした嘘でつなごうとした。僕はひょっとしたらどこかの大学の教授になって、ひょっとしたら研究所も立ててもらって所長兼務になるかもしれないけど、そのときは君僕のところに助手で来てくれるかなあとは言わなかった。ちゃんと人事秘を守りました。仮に小保方さんがそのことを誰かに喋ってそれが他者に漏れて入札でトラブってと連なって行くと関係している人々全部の人事に影響します。小保方さんが大和さんや常田さんら自分の先生方に相談するということはあり得ますからね。
当たり前だけどとてもまじめな人です。その彼が、こう書いてるんでしょ。
>>
(2) Extended Data Fig. 7d in the Article and Extended Data Fig. 1a in the Letter are different images of the same embryo and not, as indicated in the legends, a diploid chimaera embryo and tetraploid chimaera embryo.桂氏は間違えたかもしれません。 「2N キメラか 4N キメラかは、論文の重要な論点とは考えられず、過失による可能性が高 いと判断した。」って何を言ってるんでしょ。クローンだよ。2N キメラか 4N キメラかと読んだのは桂氏だ。でも実際にはどちらでも無いと書いてある。「研究の価値を高めるために強引に胎盤と断定した可能性がある」んなら捏造ということになる。馬鹿なこと言うな。とは言え、「4N キメラ胚であることは、マウス胚撮影に用いた PC に残存する写真(2011 年 11 月 28 日撮影)と若山氏の実験ノートから確認できた。」んです。移植したドナーが違うんだよという若山さんのフロイト的告白を聞き分けろと言っても、それはちょっと無理かもしれません。
(矛盾)もう一つとんでもないことがあります。(Extended Data Fig. 7d in the Article and Extended Data Fig. 1a in the Letter are different images of the same embryo)と書いている。Extended Data Fig. 1aは(「129/Sv×B6GFP」 が正しい)んでしょ。並べて比較してみましょう。
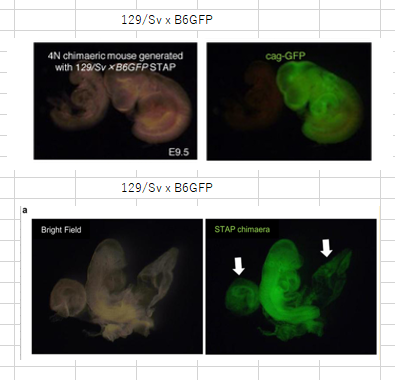
帝王切開して取り出してる一回限りの胎児なんでしょ。同じ胎児でどうして大きさと形が全然違うということがあるのですか。そっくりなのはむしろ以下でしょう。
でも、こちらはマウス背景が違うし、形も似てるがよく見ると異なっている。そもそも妊娠後同じ日数経過したマウス胎児の形はほとんど同じです。切開して広げて撮影するときの状況が少しずつ違うだけだ。Article Extended Data Fig.7d とLetter Extended Data Fig.1a は全く違うものです。
手記には「スフェアからのキメラマウスの胎盤だというものをいくつか渡され」とあるのみで、胎児はどうであったか、卵黄嚢がどうであったか、撮影した写真がどうであったかはわかりません。小保方さんが胎盤を貰ったときにマウス写真をもってなかったら何かつけるということになります。
聞けばいいだけですよ。小保方さんは7月からずっと理研にいました。実験してないときは相沢さんの部屋で待機している。相沢さんに聞いてもらえばいいだけだ。こんな明らかな矛盾にさえ気づいていないようですから、どうにもなりません。
(小保方さんのマウス背景認識)どこに事実があるのか。
まず「4N キメラ胚であることは、マウス胚撮影に用いた PC に残存する写真(2011 年 11 月 28 日撮影)と若山氏の実験ノートから確認できた。」ということです。PCに写真があってLetter Extended Data Figure 7-aの写真と全く同じであったと言ってることになるが、今見た通り別のものだ。実験ノートにマウス背景は129/Sv×B6GFPと書かれていたんでしょ。対して論文にはこのキメラはB6GFP x 129/Sv背景であると書かれ、手記によれば2012年の3,4月頃に渡されたはずの、胎盤の免染写真がセットでつけられている。以下です。
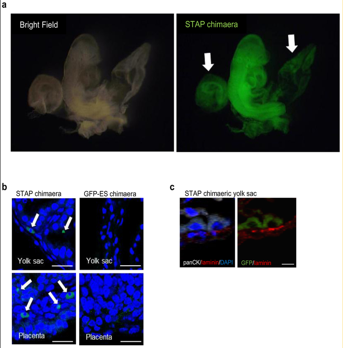
リジェンドです。
>>
a, Chimaeric mouse with STAP cells derived from CD45+ cells of B6GFP × 129/Sv mice (B6GFP, C57BL/6 line with cag-gfp transgene). Arrows indicate a placenta and a yolk sac.
b, Cross-sections of yolk sac (top) and placenta (bottom). GFP-positive cells (arrows) were seen only in yolk sac and placenta of the STAP cell chimaera. Scale bars, 50 μm.
c, Co-immunostaining showed that these GFP-positive cells (right) were found in the extra-embryonic endoderm-derived epithelial cells (pan-cytokeratin+ and overlying laminin+ basement membrane; left) of the yolk sac. Scale bar, 10 μm.B6GFP × 129/Sv mice (B6GFP, C57BL/6 line with cag-gfp transgene)と書かれています。小保方さんは親の雌雄の書き分けルールを知ってます。小保方さんはB6側にCAGが入っていると聞かされているんです。Acrに関してはそもそも小保方さんの実験目的には関係がないから知らされてないだけで、実際にここで使われているのは岡部マウスのB6GFPなんです。Article Extended Data Figure 7のマウス背景の書き分け方を見ておいてください。前がメスと知らないでこんな書き分け方をするわけがありません。
>>
B6GFP x DBA/2
129/Sv x B6GFP
Letter Extended Data Figure 7-aのマウス背景記述を小保方さんのマウス背景の書き分け方の不注意と書いたのは桂報告書で、小保方さんに確認したわけではない。
>>
なお、図の説明にある「B6GFP×129/Sv」 は、最初にメス、その後でオスの遺伝的背景を書く通常の表記法では「129/Sv×B6GFP」 が正しいが、不注意による間違いと思われる。 小保方さんは2012年3,4月に免染依頼を受けた胎盤のマウス背景をB6GFP×129/Svと聞いていたからそう書いているので、それを2011/11/28のキメラが129/Sv×B6GFPなんだからときめつけて、不注意による間違いだと判断する前に、本人に確認すべきです。7月からずっと再現実験で理研に来ているんですから、聞いたら分かることです。
因みに彼女が不注意な書き方をしているのはむしろArticle Figure 4-cの以下の部分でしょう。
以下がそのリジェンドですが図のキャプションと違って、逆になっている。これは2月の分ですからリジェンドの方を間違えている。
>>
c, Adult chimaeric mice generated by STAP-cell (B6GFP × 129/Sv; agouti) injection into blastocysts (ICR strain; albino). Asterisk indicates a highly contributed chimaeric mouse.恐らく、以下のFigure 4-cのリジェンド背景記述と混同したのではないでしょうかね。
リジェンドは以下です。これは論文に添付されているヴィデオのストップモーション画像だとされています。無論、若山研で当時撮影されていたものです。笹井さんのところではキメラがないから撮影はできません。
>>
f, E10.5 embryo generated in the tetraploid complementation assay with STAP cells (B6GFP × 129/Sv).もう一つ紛らわしい書き方をしているのはアーティクル本文の以下でしょうか。ここでは雌雄を配慮せずに書いている。
>>
Furthermore, in a tetraploid (4N) complementation assay, which is considered to be the most rigorous test for developmental potency34, 35 (Fig. 4a, bottom), CD45+ cell-derived STAP cells (from F1 mice of B6GFP × 129/Sv or DBA/2) generated all-GFP+ embryos on embryonic day (E)10.5 (Fig. 4f, Extended Data Fig. 7d and Supplementary Video 3), demonstrating that STAP cells alone are sufficient to construct an entire embryonic structure. 以上の検討からわかることは2011/11/28の写真が本当にLetter Extended Data Figure 7-aの写真と同じであったのなら、小保方さんが写真だけを選択し間違えたということになる。そして、同時に小保方さんはその写真を持っていたということです。2011年の胎盤の免染なんてあるわけがないと我々が知っているのは、手記を読んで胎盤の免染がいつの話か読んでいるからです。桂チームは無論手記は読んでない。若山さんに聞いただけですね。2012/11/28の胎盤を分析したなんて聞いてないんじゃないですか。若山さんがそんなことを言うはずもない。キメラが初めて成功したというのに、胎盤が光っているかなんて誰も気づきませんね。そして胎盤は生ものです。その時のものだ。免染写真が2011/11/28のキメラのものであるわけもないことです。だとしたらただの写真の選択間違いに過ぎない。では胎盤を免染させた時のキメラ写真はどれだと調べて行かないといけないですよね。ところが彼らの結論は以下でしたね。
>>
2N キメラか 4N キメラかは、論文の重要な論点とは考えられず、過失による可能性が高 いと判断した。STAP 細胞の胎盤への寄与は、Letter の論点として重要であり、研究の価 値を高めるために強引に胎盤と断定した可能性があるが、調査により得られた証拠に基 づき認定する限り、研究不正とは認められない。小保方さんに確認したらすぐにわかることを確認もせず、ただ若山さんに言を左右にされて結局分からなかったということでしょうが。
小保方酸浴細胞核使用ntESキメラだと気付かない限りは全部を整合的に理解することはできません。この時点で桂チームがそのことに少しでも気づいていたということはなさそうです。変だなあとはだれしも思うでしょう。でもど素人の私は分かるまで5年かかった。専門家はもっと早く分かるはずですが、こちらの方は、その時にはすでに文科省の圧力がかかって緘口令が敷かれているんでしょうね。
(相沢さんの感想)ずっと以前にOoboeさんのパートナー氏が何かの序に相沢さんに聞いてくれたことがあります。ntES化したと言ってる人がいますけどと。言下の答えは、それだとどちらの性質なのかが分からない、というものでした。
ntESはキメラができます。従って小保方細胞をntES化したらキメラができるのは当たり前です。相沢さんはその時よく考えずに、やはりキメラができたことがSTAP細胞の多能性証明なのだから、そんなことしてキメラを作っても無意味でしょと言うコンテクストに沿って反論されたんでしょうね。特に彼が直前まで携わっていた検証は小保方細胞でキメラが再現できるかという仕事でした。ここにntES論を持ち込まれたら誰でも意味が分からないでしょうね。
理研は特許を申請しています。その趣旨は論文の記載に沿ったものだ。論文に書かれている通りの現象が再現できなかったら特許申請を維持していると大変な損害を被る。できなければ取り下げなければならない。今はどうかは知りませんが、当時は論文のコンテクストに沿って現実との乖離を理解しようとしていたことは当然です。
でも我々のntES論はそういうものではありません。若山さんは何度もキメラ作成を試みてできないことを確認している。恐らく写真にあるようなナイフ切り分けでもできないことを確認しているはずです。
小保方さんの細胞はキメラのできない細胞なんです。若山さんにとってそれは科学的事実です。相沢さんは論文に沿って考えているし、それが仕事ですから、キメラは出来たと書いてあるというところから離れることはできません。でも、若山さんは2011年の10月頃にできないと確認し終わっているんです。我々はその若山さんの確信に沿って彼が何をしたかを考えている。論文なんてこの頃書かれていない。過去を知りたかったら過去の人になり切らないといけない。笹井さんの論文なんて存在していません。ここにあるのは若山さんの意思だけです。
小保方さんの細胞はOct4-GFPを大量に発現しています。後にはノフラー氏とか関さんが自家蛍光だなんて宣伝をしまくって世間を過った方向に連れて行こうとしましたが、ど素人じゃあるまいし、電子顕微鏡の自家蛍光識別もしない専門家がどこにいるでしょうね。顕微鏡下でよく光ってるねと言った若山さんがフィルター切り替えすらしなかったと馬鹿にしているのでしょうかね。それとも自家蛍光は6時間程度しか続かず、ずっと光ってることの分かるライブセルイメージング実験が最初若山研で行われているのだということすら聞いてなかったのか。
今、丹羽さんの論文でGOFマウスのGFP漏れ出しが報告されました。これは内在性Oct-4が発現していないのにOct4-GFPが発現する現象です。丹羽さんの検証論文以前には誰も知らなかった科学的事実です。自家蛍光だ、酸浴させると内在性Oct4が発現することもあるのだなどという机上の空想を述べる人はいましたが、誰一人、GFPだけが発現するなんて事態を予想した人はいません。小さな新事実ですが丹羽さんの発見なんですね。
このことの意味を論じる人は知る限りありませんが、GOFマウスを開発した人々にとっては研究課題になってるはずですがね。通常の使い方ではGOFマウスは内在性Oct4を発現させるプロモーターが働いた時に同時に同じプロモーター下にGFPを発現するように設計されている。酸浴させ、亜致死下に置いたら、GFPだけが発現するなんて初めての知見です。
歴史的に考察するというのは、この知見は今は知られているが、当時は知られていなかったという地点に想像力を働かせて戻れるかということですね。関さんもノフラーも知らなかったことです。知らなければ本物か自家蛍光かしか推測ができない。彼らにとっては丹羽さんの発見は思いもよらないことでしたね。専門家だから専門領域に関する主張が正しいなんて保証はありませんね。特に自然科学では知られていないことの方が多いんですから当然です。専門家は現時点で分かっていることの範囲内で解説ができるということだけです。
重要なことはGFPの漏れ出しなどという現象があるかもしれないなどとは若山さんも、小保方さんもこの時点では知りません。「よく光ってるね」と言ったんです。それは自家蛍光だよとは言わなかった。その代わり、6時間で消える自家蛍光を取り除くために一週間に渡るライブセルイメージングを撮影した。若山さんは自家蛍光なんてこの頃考えてもいませんね。この実験は笹井研での映像以前に若山研で行われていることが手記に書かれている。手記も読まないでこの事件に関して何かを語るなんてをこのことですね。関連する文献は全部読まないといけない。そして眼光紙背に徹してからものをいうものです。
若山さんがこの細胞は一体何なのかと考えたとしたらそれは何か奇妙なことでしょうかね。後の論文を読んでそのコンテクストの上でしか物事を見れない不自由さを背負っていては歴史は見えないでしょう。後の論文は後にそうなったので、この時には姿もないものです。あるのは若山さんの好奇心だけですね。
彼は小保方さんのためにもう少しこの細胞の性質を見極めてやろうとしたんでしょう。無論、彼自身の好奇心もあった上に、彼は小保方さんをヴァカンティの手元から奪いたかった。いたく気に入ったんですね。4月に腰かけて小保方さんが渡米できるようになって帰ろうとしたときに自分のラボに誘った。手記も読まないでどうしますか。手記の記述は小保方さんがその後なぜ若山研の客員になったのかをちゃんと説明しています。若山さんが誘ったからこういうことになった。誘わなかったら彼女は米国で研究していたでしょう。こういう裏事情も知らずにこの事件を語ろうなんて愚かしい。
彼女はキメラができなかったのでヴァカンティの許に帰ろうとした。できなかったからESで捏造するなんて、Oct4-GFPがあんなに光っているのにそんなもったいないことをする研究者はいないでしょう。ティシュー論文や、博論段階では何十個のスフィアに一つという細胞であったのに、若山さんのところに来て酸浴実験をしたらどんどんできてきたと思い込んでいる。本物と信じ込んでいるGFP蛍光を見ているのにどうしてそれを捏造で台無しにするなんてことがありますか。
この点に関しては若山さんも同じです。若山さんもGFPの漏れ出しなどというアーティファクトは知りません。内在性Oct4を発現する細胞がたくさんできてきていると思っているんです。
小保方さんは渡米してその細胞の性質を見極めようとした。若山さんはちょっと待ってと引き留めたかったんです。米国に帰らずにもう少しここでやりなさいと。
相沢さんはntES論をあの時点では一蹴しました。今はどうか。いまでもこの問題は根が深いと思われますね。一度自分のntESの材料にしてみようと思いつくのはそれ程むつかしくはない。できてきたキメラが普通のntESとどう違うかを調べてみようとする発想です。でもここには相沢さんの言った、方法論上の問題がありますよね。論文はキメラができたことを主張している。その論文から今離れるわけですから、相沢さんが一蹴した根拠はそれなら消えているかというと、消えません。特に最終的には胎盤貢献が違うのではということになったんだけれども、ではコントロールとしての普通のntESの胎盤は既に解明しつくされているのかという問題です。ntESの胎盤異常はとても有名な研究課題です。どちらもはっきりしないような問題ではコントロールが取れないではないかという意味においてなら、相沢さんの指摘は正しいでしょうね。そしてそれが若山さんの致命的な誤認の原因になった。
こういう風に推測してみる。
①小保方さんを山梨に連れて行きたかった。
②GOFのGFP漏れ出しは当時誰も知らなかった。若山さんも小保方さんもたくさんできてきていると誤解した。
③若山さんは小保方細胞の性質を調べるためにその核を使ってntES化した。
④違いは胎盤貢献ではないかと思い込んだ。
⑤しかし、胎盤貢献に関してはコントロールとしてのntES側がまだ研究途上で確定事実がなかった。
⑥②の誤解からそのntES化はただのCD45陽性細胞の核移植結果となってしまっていたが、若山さんは気づいてなかった。
⑦結論的には彼の小保方細胞核を使ったと信じていた現象は只単に自分のntESの性格であったに過ぎなかった可能性がある。もしそうなら自分でやればいい研究だったということになる。
⑧しかし、丹羽論文が出る前にそのことに気づくことはできない。彼は彼なりに自分の研究にたくさんの疑問を抱いていたはずである。
⑨スタンダートなプロトコルでキメラが出来てないことは彼がリクルートのためについている軽い嘘にすぎなかったが、自分の行っている研究は本物であって、自分の本当の研究を隠すために嘘をつき続けていても後のイクスキューズはできる。
⑩しかし、自分の行っている研究に何か勘違いがあったら、それは小保方さんに対してついている嘘のイクスキューズをも失ってしまうことになる。
⑪何も知らない笹井さんが若山さんの研究成果を使って査読対応実験で調べたレター論文の実験は彼を震撼させたかもしれない。
⑫若山さんは2013年の8月にレター論文の原稿を読んで、自分でも分からない論文になってしまっていたと、笹井さんに責任著者を降りたいと申し出た。何か自分の間違いに気づいたかもしれない。それは相沢さんの言った方法論上の瑕疵だったかもしれない。
他方、遡って、ヴァカンティは論文が出るまでは自分のところから小保方さんを離さないと言った。では論文は書かせないといけない。あちこちにぶら下がったまま忘れられていく論文が大半なんですから、取り敢えず最終的にはヴァカンティの主催のティシュー論文でぶら下がったままにすればいい。
案の定、ネイチャーはリジェクトした。だったら次はティシュー誌に挙げて置いたら約束は果たされて、小保方さんは自分と一緒に山梨大に来てくれるので無いといけない。約束じゃないか。
でも、デイナのニューヨーカーへの寄稿文によると、ヴァカンティは低いバーを飛ぶつもりかと小保方さんを励ました。なぜか。キメラが出来ていると嘘をついているままだからだ。キメラが出来ているのにノーベル賞級の発見で無くてなんだ。ヴァカンティでなくても誰でもそう思いますよね。若山さんの誤算ですよね。薬が効きすぎてしまってる。でも、歴史的視点を失わないでいればこのときここで行われている実験を世界中で知ってるものは10本の指でしか数えられないほどの狭い範囲だった。それが先進国中に知れ渡るほどの問題に発展するとこの時点で予測した人はいたでしょうかね。ヴァカンティさえごまかせたらなんか小さく丸め込めそうでもある。いざとなったらああ、あれはESコンタミだったみたいと。
C. 129/Sv carrying Rosa26-gfpa. AC129の実験とTCR再構成調査以外の細胞追跡法(「僕のマウス」証言)所謂「僕のマウス」が登場するのは2014/6/16の若山さんの記者会見ですね。「僕のマウス」を渡したと証言した。ところがそういうマウスを渡したという実験ノート等の記載証拠は全く無いどころか、論文には明らかにB6側のGFPヘテロ記載のマウスしかない。実際の実験を担当した共著者である若山さんが論文にヘテロだと書いてあるのに今まで一度も小保方さんに注意していない。今頃何を言い出すのだというトンデモ主張である。それを桂報告は確認もできないまま諸幹細胞等からGFPヘテロが出たからと言って、小保方さんが太田ES等を使って捏造したというストーリーにしてしまったのである。警察もまま無理な立件をすることもあるが、これほどの馬鹿なことはしないですね。「僕のマウス」渡したんですと主張したら、はい、その記録を見せてくださいとまずは尋問過程で事実を確定させていく。この桂報告書は最初の時点から証拠無しの前提なのである。それどころか論文にヘテロと書いてあることの経緯が何かということすら調査していない。
これはよほどのことですよね。万が一殺人事件に巻き込まれて警察にこんなことされたら誰でも殺人犯にされてしまう。無論、事件の裁判なら弁護側と検察側で証拠の能力検証が行われるので、少なくとも事実認証に関してこんなことはありえないが。この事件は裁判ではない。第三者委員会が調査した調査結果なのである。
「僕のマウス」を渡したんです。ノートの記載はしていないが僕を信じてください。論文に小保方さんがそういう記載をしていることは知らなかった。彼女はマウスのことは詳しくないんです。という感じで、ああそうですかとなった。小保方さんに、確認もしていない。ホモかヘテロかは外見では見分けがつかない。論文になぜヘテロ表記したのですかと聞いたらそういわれたと答えたと思いますね。マウスに詳しくない人は言われたとおりに書くのが普通ですね。最初の成功キメラはGFPヘテロのF1でしたよね。それは実験ノートに書いてあった。それが分かっているのに二回目が「僕のマウス」であることはそのまま信じた。
「僕のマウス」を渡したのにヘテロマウスが返ってきたと主張しているのですから、何より大事なのは「僕のマウス」を渡したという証拠です。最初の成功マウスはヘテロマウスを渡したらヘテロマウスが返ってきてるんですから、二回目もそうじゃないのかねとすら問うてない、というより気が付きもしていない。
(「僕のマウス」の初出)所謂「僕のマウス」は記者会見で出たものですが、このマウスらしきものの初出は2012年の5月頃です。若山さんはFLSのキメラを作ってそのキメラのジャームライントランスミッション確認実験を行った。キメラは20日で生まれますが、子供を産めるように成熟するまで50日かかる。その後交配して20日後にキメラ子が生まれる。キメラ移植からほぼ90日後です。
FLSの実験は1/31にFLS1、2/2にFLS2~8の二日間行われた。小保方さんは1/23に初出勤している。それまではハーヴァードで仕事をしていた。以下謝金票の再掲です。
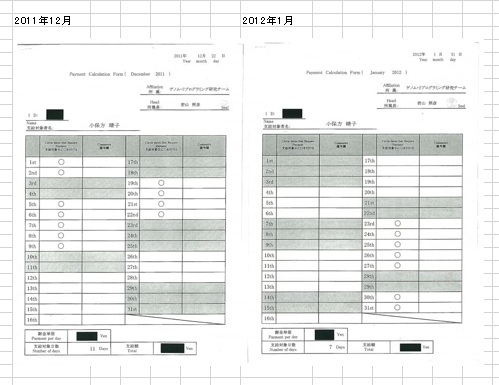
若山さんが後にあれは「僕のマウス」だったと言い出した赤ちゃんマウスを数匹渡されたのは出勤日かその翌日ですね。インキュベーターで1週間かかりますからね。研究者は土日無しみたいなものですからこのときの出勤がどうであったかは分からないが、ぴったり1/31の開始に間に合うようにすべては段取りされていたということです。
因みに彼女が12/27テラトーマを切開したのは1/24だと証言されています。4週間後です。彼女はティシュー論文と博論は6週間で切り出し、アーティクル論文では博論のを使ったから博論の時のままの記述で、6週間でかつNOD/SCIDだと書いている。ヌードマウスではない。永田龍平衆議院議員が指摘しましたね。12/27テラトーマはヌードマウスでの実験で、4週間後の切開ですが、HE染色しただけで免染しなかった。おかしいと思ったからです。後に調査の時小保方さんはこのおかしいと思ったということを言い出せなかった。誰かを疑うことになるからですね。これが言い出せなかったから写真を間違えたと嘘をついたんです。それで話がこじれた。しかし、調査チームが既に偏向していますからね。彼女は本当にキメラができたと信じていますから、そういうことは言わなくても真実は明らかになると思ってたでしょうね。
若山さんは小保方さんが博論の写真を使ってたと聞いてあり得ないと思い態度を翻したと証言していますが、テラトーマの上からできたばかりの幹細胞を注射したのは自分ですから、小保方さんがES混入を疑ってあのテラトーマを使っていなかったのだと知ってショックだったでしょうね。あのころから疑われていたんだと。山梨大に助手で来ないかと誘われて、即答できなかった事情とも関係あるかもしれない。
ともあれ、幹細胞は若山さんは3,4回の植え継ぎで樹立判断をしているようです。2週間程度の後でしょうか。FLSの樹立です。ここから幹細胞のキメラ実験に入る。
その日を2/15だと設定すればその90日後は5/15だ。5/15前後に若山さんは小保方さんにGFPが半分にしか来なかったと言った。それはどの実験なのか。
2Nキメラはリシピエントの胚由来のキメラも生まれますから、これらを兄妹交配するとGFPが来たものが分かる。一つの株でたくさんのキメラ胚を作るので、キメラもたくさん生まれる。その一つにでもGFPが来たらジャームライントランスミッションは有りですね。無いことを証明するためにはすべての可能性を虱潰しにしないといけませんが、有ることの証明は一つあったらいいんですね。
以下が2Nの結果でした。全ラインyesですね。
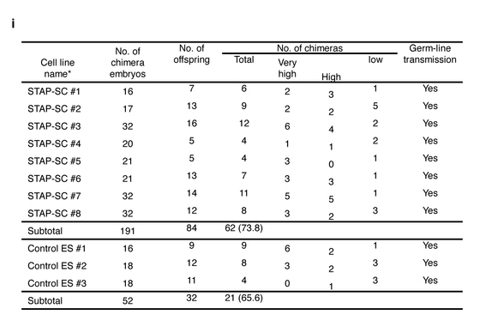
若山さんは全部に来ないといけなかったのに半分にしか来なかったと言った。これも不可解な発言ですよね。これは2Nの実験ではあり得ませんね。雌雄ともに配偶子がリシピエントのキメラも含まれているわけですから全部に来るというケースはない。4Nの実験ですよね。
以下がそれです。
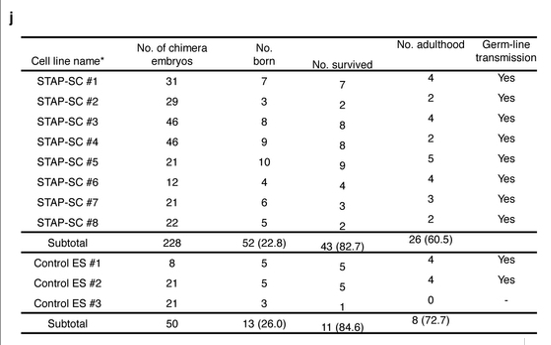
4Nキメラは全身がドナー細胞由来ですから配偶子もドナーです。この配偶子に生殖能力があるか無いかを確かめるのがジャームライントランスミッション実験です。では2Nと同じく兄妹交配させればいい。ところがここには書いてないが228個のキメラ胚から生まれてきたキメラ52匹の全てが雄だったんでしょ。この実験は無論若山さんが一人で行ってデータだけを小保方さんに後から渡しているものですね。
このSTAP-SC#1~#8ってFLS1~8のことですね。これは若山さんが放医研に調査に出して全ライン雄と発表したものですよね。雌が居ないんだか4Nキメラ子同士の兄妹交配はさせられません。既述していることですね。では一体メスはどこから持ってきたのか。
桂報告書によると若山さんは戻し交配したと言ってます。通常戻し交配というのは実際の親に掛け戻すことです。彼がロックフェラー大学で作っていたB6のCAG-GFPホモマウスからそのGFPだけを129/Sv(GFP無し)に移すときに使ったような手法が戻し交配です。若山さんはこの時のドナーは「僕のマウス」を渡したと言ってるんですから、本当に"戻し交配"なのなら雌の129/Sv-CAGホモを持ってくることになる。
でも、親にCAG-GFPホモのを持ってきたらキメラにジャームライントランスミッションがあったのかどうかはGFP蛍光確認では調べられませんね。全部光ってしまいます。
ジャームライントランスミッションの確認のためには、CAG-GFPの無い、しかし、生殖能力がちゃんとあると分かっているメスを持ってこないといけない。で、CAG-GFPの無い129/Svを持ってきたということですが、この場合"戻し交配"とは言いませんね。これはワイルドタイプの無関係な背景のメスでもいいわけです。この場合だとはっきり"戻し交配"と言わないことは分かりますね。同様にGFP無しの129/Svのメスであっても"戻し交配"とは通常は言わない。この言葉は調査に対する尋問への言い訳対応の中で出てきた言葉です。小保方さんに対する説明時には無いものですね。
228個のキメラ胚移植の結果52匹の4Nキメラが生まれてきたがそのすべてがオスだった。彼はそれを不思議にも思わずに、4Nキメラ子にメスが一匹も居なかったので、キメラ子のジャームライントランスミッション確認実験をするときに、GFPの無い129/Svのメスを使ったと説明していることになるんです。常識的には全部オスだということが彼にとって何も不思議ではなかったから、実験を続けているだけでしょう。
我々のntES論ではFLSの元となったたくさんある中の一つのntESがたまたまオスだったもので、それを8株に分けたものがFLSとして小保方さんに渡されたものですから、GFPがヘテロなのは若山さんには分かり切っていたことだ。そのジャームライントランスミッション実験に129/Svを持ってきたら"戻し交配"だということになる。なぜなら、若山さんがntES化させるために小保方さんに作らせたSTAP細胞の材料は129/Sv x B6-CAG-GFP(岡部マウス)だからです。彼は「僕のマウス」を渡したということが嘘であることを"戻し交配"という言葉を使った時に苦し紛れに告白してしまっているんです。
実際に実験に使われたのは成熟した26匹ですね。4Nキメラで、しかも全部オスですね。若山さんが渡しているマウスは129 x B6-GFPです。精原細胞から減数分裂で129とB6GFPの二種の精子に分かれる。ここに129のメスを持ってくると、GFPの有るものと無いものと半々に生まれてくる。当たり前です。上表では全部Yesになっていますが、キメラは最低2匹いますから確率的には全部Yesでもおかしくはない。2匹のうちの1匹でも光ればYesです。
実験そのものは岡部マウスとのF1での小保方細胞核使用ntES化の実験のシリーズとして何もおかしくないものです。
ヴァカンティ氏が米国特許仮申請をした頃に、若山さんが、小保方さんに半々にしか来なかったのが変なんだと吹き込んだ。それはヴァカンティ氏に対してあれは何かの間違いだったという言い訳のための伏線だったんですね。何か、マウスの交配ミスがあったのだという言い訳です。この頃まだ理研でのネイチャー論文なんて書かれていません。このFLSのキメラ実験なんて三誌論文には無いものです。
ネイチャーはリジェクトされたんだからヴァカンティさんは小保方さんの論文をティシュー誌に載せて置いて、一年後に進展が無ければ仮申請は引っ込めたらいいんです。小保方さんは山梨に連れて行っていいんでしょ。そういう約束だったじゃないか。
他方で、ntES化の実験結果はまだ若山さんを魅了している。若山さんが何かこの研究に関して捏造を行うということは常識的にはあり得ません。彼は既に功成り名遂げた学者でノーベル賞を待ってる人なのでそんなことをして得が無い。又何か大発見したんじゃないかとワクワクしてるだけです。小保方さんと一緒に若山研でもう一度何か出来るかもしれないと夢見てる。
ここに何かしら、GFPの漏れ出しなんかも含めて、いろいろと大きな勘違いがあったことと、もう一つは小保方さんのリクルート絡みでヴァカンティ氏に対して策を弄している。これが思いもかけない理研の参入という事態に至って、若山さんは窮してしまったんですね。逃げ場がない。論文が落ちてくれることを願いつつ、通ったらどうしようかと考える。それが太田FESのコンタミストーリーです。
(小保方さんの+/-)専門家の間ではヘテロ表示を+/-と略記する。小保方さんが自分のサンプルに+/-と書いているものが以下です。木星リスト103,109,111,114番です。

小保方さんは若山さんからこの時に本来ならGFPが全部に来るはずのマウスを君に渡したんだぜと刷り込まれているんです。だから、若山さんが記者会見で突然「僕のマウス」を渡したと言われて心当たりがあったからこの件に対して反論できなかったんです。そういえばあの時そんなこと言ってたわと。でも違う細胞が返ってきたということに関しては否定しましたね。渡されたマウスで作った細胞をお返ししたと。
因みにES+/-は後にそう命名されたFLSのことだとはっきり分かりますね。小保方さんは5月頃に若山さんにそう言われて後に再凍結したラベルに+/-表示を加えているんです。ESは学生にもらってGOF-ESしか持っていませんから、ES-likeの意味でESと書いていた。129 GFP +/-も同じですね。彼女F1はB6にGFPのあるヘテロだと思っていますから、129GFPと書いたら129B6と書いているのと同じになる。当時紛らわしいものがなかったからこう書いている。129F1GFPは紛らわしくなりかかっていた時期かもしれませんね。仮に129/SvのGFP入りの実験が始まると以前のFLSはF1と入れておかないと識別できなくなる。この114番はその2となっていてその1が無い。たぶん件の95番がそうではないかと思いますが、先に2を凍結して、これを1に書き換えるつもりが渡米するときの多忙でそのまま以前のラベルで再凍結したものかもしれないが、その辺りは分からないですね。ただ、そもそもヘテロだと思っているところにホモだった筈なのにヘテロだったよと言われて、最初からヘテロだと思っていたけどやっぱりヘテロだったんだという意味で+/-表示しているんですね。本来なら書かなくてもいいくらいの情報だ。まんまと刷り込まれている。
若山さんは3/5前後には生まれたキメラが全部雄だと知っていた。そしてキメラが成長するのを待って5/15前後にジャームライントランスミッション結果を知らせるときに、「僕のマウス」の情報を小保方さんに刷り込んだ。
小保方さんの最初のネイチャー投稿論文のリジェクトは4月中です。まだジャーライントランスミッション実験の最中でしょう。そしてヴァカンティが米国特許仮申請をしたのが4/24です。
ここに「僕のマウス」ESの樹立開始日が2012/4/19である謎が重なる。これは件の129B6F1ES-1です。これによってAC129とFLS-Tが捏造されている。若山さんはMTAの添付資料リストにこの樹立開始日を2012/5/25と書いていて1か月強の差がある。
この頃又胎盤が光るといわれて胎児胎盤を渡される経緯もあって、このキメラの背景は明確には誰も語っていない。
更に4月のネイチャーリジェクトを受けて西川さんのTCRアドヴァイスがあった。CD45とSTAP細胞からは簡単にTCR再構成の全バンドがでた。しかし、幹細胞に関しては若山さんはテクニカルスタッフの二人の内の一人に任せ、小保方さんにやらせなかった。結果は全バンドは出てないという意味では無かった。小保方さんがやりなおしても無かった。そんな筈はない。ただSTAP細胞を培養しただけのポリクローナルな細胞集団であるはずだ。
出来事の目白押しした時期ですね。そしてAC129の実験が始まった。
実はntESキメラを作るのには事前にクローン胚に入れて3.5日の胚盤胞を取り出した後に中のインナーセルマスを取り出してES樹立します。一週間から半月位多くかかります。5/15は5/末から6月初頭である可能性まで考えておかなければなりません。
- 2019/09/06(金) 20:48:34|
- AC129
-
-
| コメント:0
d. TCR検証との関係(西川氏のアドヴァイス)AC129の実験が「遺伝的背景が及ぼす影響」を調べるためでなく、細胞追跡の実験だったらどうでしょうか。その可能性も念頭に置いて検討してみましょう。そうでないと、なぜこの実験に関して129ローザなんて話が紛れ込んでくるのかが分からない。何かまだ知られていないことがあるからとんでもない"勘違い"だという説明になってしまう。誰がローザと聞きもしないでローザと書くでしょう。誰かがローザと言ったからローザと書かれている。それを追求しないと、単に勘違いで済む問題ではありません。自分に置き換えてみたらわかる。聞きもしないで、かつ何の理由もなく、突然ローザと書けるか。はは。
我々の仮説では、若山さんは小保方さんの細胞がキメラ形成しないことは知っています。しかし、何物かではあると信じて、或は何物であろうかということを知ろうとして、小保方酸浴細胞核を使用してそのntESからキメラを作ってみている。そしてどうもその胎盤が光っているのではないかと気づいたわけです。だからこそそのntESをTS培地によって誘導してCTSを作ろうとした。
ただし、この胎盤蛍光が単なる希望的思い込みでなかったかということが後の議論でも出て、かつ小保方さんの免染も確かにGFPは入っているんですが正しくその場所を免染しているとは限らない。つまり光って当たり前の場所を調べてたのではどうしようもないという疑義がでたわけです。このことは笹井さんが参加してからのことで、査読者たちに指摘されたり、事件発覚後にパブピアで指摘されたことなので、この当時は論文も幹細胞化の実験に関しては発表していないから、胎盤の件が正しいか否かは二人ともまだはっきりとは分かってないんです。
一方でACCsの論文がネイチャーにリジェクトされ、若山さんにしてみたらリクルート絡みの問題は終わりだと思ってたら、ヴァカンティが高いハードルを飛べと小保方さんに指示したもので、論文はティシュー誌でなくセルとサイエンスに投稿することになった。
どうしたら通るかという小保方さんの相談を聞いてもらうために若山さんが西川さんのところに連れて行った結果、TCRアドヴァイスを得た。そして実験して結果を添付したがまたセル、サイエンスともにリジェクトされた。
(PCRの結果)一方でFI幹細胞の研究をつづけながら、若山さんは小保方さんからのTCR結果を聞かされている。彼は小保方さんの作ったリンパ球の酸浴細胞にTCR再構成PCR結果が出るのは当然だと思ってる。CD45+だけでも出るものを念を入れてFACS選別で確率高くT細胞だけに絞ってます。ある程度違う細胞が混じってもPCR検査では出るに決まっているし、現に出てる。もう1度貼り付けましょう。
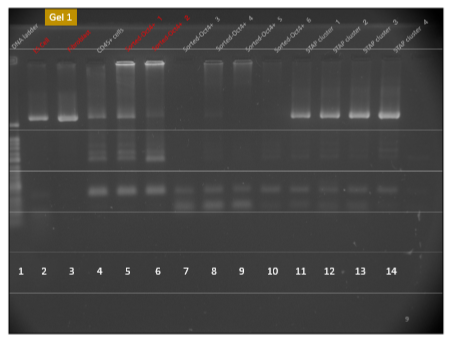
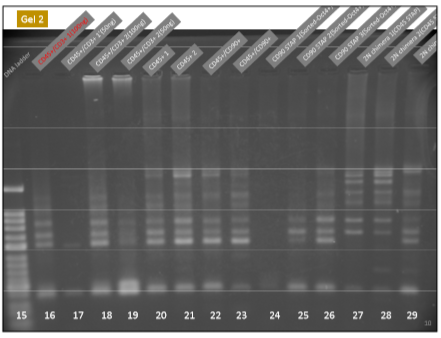
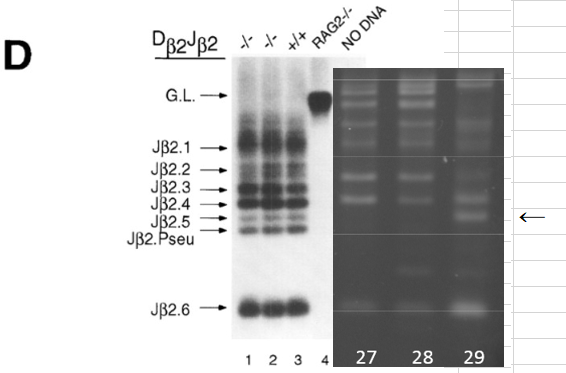
ど素人なんでどれがTCR再構成バンドか分からなかったから苦労しましたが以下が泥縄のお勉強の成果でした。
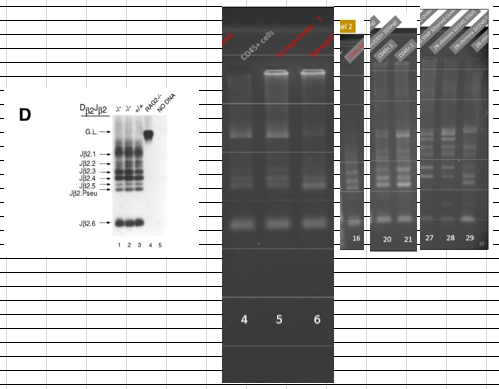
はっきり出てますね。CD45陽性細胞と酸浴STAP細胞にはTCR再構成はあります。ここは問題ない。キメラにもTCR再構成があるが、ここはどういう出方をしているかが問題になった。そしてGel1とGel2には所謂STAP幹細胞のPCR結果がありませんが、幹細胞は別のラボメンバーが担当した。これが後に問題になった。
ここで残されている課題は
キメラと幹細胞にはどういうバンドが出ているべきなのかという問題です。
STAP幹細胞は若山さんが小保方さんから渡された細胞を基本ES培地で培養誘導して作られたと若山さんが語っているものを小保方さんが論文にプロトコルとして書いている。手記によれば小保方さんは自分ではプロトコルに従っては作れていない。作れないのに構わずに書かせたのは若山さんですが、丹羽さんも相談を受けて若山さんを信じるべきだと答えている。こんな論文はおかしいと誰でも思う。でも、笹井さんと丹羽さんは業務命令でリヴァイズの手伝いをしていますからね。実験が疑わしいというスタンスは初めからありません。
筆頭著者ができないのに責任著者が自分はできるから、できたと書いて置けと言われ、シニアの指導者も若山さんを信じるべきだと助言した。後から、若山さんはあれは小保方さんがESを自分に渡したのだと言い出した。丹羽さんは梯子を外されたんやと、その足で山梨に向かったという。記者会見でなにしろESを渡されたと思い込んでいるもんでと、語っている。
ESを渡されたら誰でも気づくということは既に我々が証明した。若山さんは嘘をついている。論文は通らないようにと書かせているんですね。それが通ってしまったんで、逃げようとしたんです。通りそうだと寺下さんから情報を受けた頃からあちこちに情報リークをして準備を始めている。そしてタレコミがあって事件化するまでは知らぬ顔をして小保方論文をほめていた。11次元の博論のテラトーマ画像流用指摘から突如豹変した。予定通りです。11次元に博論の件をあらかじめ知らせたのは若山さんですね。
ここから先はかなり悪質ですね。どうしてこんなことになったか。何かやはりリクルートの経緯だけでなく、若山さんを心情的に怒らせるものが加わっているのではないかとも疑義される。事件に無関係なはずの遠藤氏の、舐めてますねこれという、やくざ的な言葉づかい、或は真綿でじわじわなんていう、会ったことも無い相手に対するいわれのない憎しみの原因がどこにあるかと推測すると、若山さんからの情報ですよね。俺達を舐めてると。どういう情報のやり取りになっているのでしょうかね。
(研究の発案と特許)若山さんは小保方さんが山梨に来てくれると思って、或は来て欲しいと強く望んで、幹細胞化の論文のためのデータを渡している。ところが小保方さんはそのデータを持ったままヴァカンティの許に帰った。
Ooboe さんとパートナー氏の公開請求で共同研究契約書は交わされていないと判明している。理研の事務窓口は共同研究の実態があったのかどうかも分からないと答えている。
すると小保方さんの客員身分はどういう手続きで生じたのか。どうやら、客員受け入れ申請書の類のものらしい。そしてその保管期限も過ぎて書類は無いという。
では客員申請はどういうケースで行われるかというと共同研究申請書に基づくのが普通であるが、今、理研のその規定は見れなくなっている。どうやら安易なイレギュラーな客員受け入れが行われていたようですね。そしてそういう受け入れは海外でもあって、学者が海外旅行のために休暇を取るのにこの契約がまま流用されているらしい。このケースでは小保方さんの震災避難の受け入れ、或はそれ以前の博論実験の受け入れでもそのイレギュラーな処理が行われているのではないかと疑われる。契約書が無いというのは正式な理研の答えで、かつ、岸氏がちゃんと約定してないからこうなると怒ったという情報も流されている。
そういう研究状況の中で、小保方さんの論文の投稿と、若山さんの幹細胞化の実験が並行して進められている。スフィア細胞自体はヴァカンティ研での小保方さんの発見であるが、理研に来て酸浴実験からGOFの赤ちゃんマウスを使ってのOct4-GFPの蛍光までの実験は若山研での発見である。しかし、約定の無いままになんだか分からない発見らしきものが次々に続いていく。
特許でもヴァカンティと揉めている。11月に小保方さんがヴァカンティの許に戻った時、幹細胞化実験のデータは小保方さんが持ち逃げした形になっているが、渡したのは若山さんで、しかも共同研究契約書は存在してない。小保方さんが若山さんに渡されたデータは一体誰のものなのか。
岸氏が約定もなくこんなことをしているからこんな問題になると怒ったらしいのは一般人でも当たり前だと思うところでしょうね。でも本当は若山さんはそのままヴァカンティ研で小保方さんが研究を続けてくれていた方がありがたかったでしょう。何しろntES化しないでキメラの再現はないし、幹細胞化の実験の意味も分からないはずだ。どうしてあの時キメラができたのか。別の実験のESをコンタミさせてしまっていたみたいと言い訳したらそれで済む。
ところが小保方さんは理研に呼び戻されてしまった。しかも笹井さんと丹羽さんがメンターになるという。とうとう本当のことを言えないままに論文が自分の手を離れてしまった。笹井さんの手にかかったら論文は通るかもしれない。キメラはスタンダードな方法でできたと書かれる。再現ができないと最後に自分のところに責任が来る。論文は通ってもらいたくない。せっかく三誌は落としてくれたのに今更別の雑誌に又投稿するなんて困る。それには幹細胞化の論文をつけさせて、一度落としてくれたネイチャーに再投稿するのがいい。こんなntES化実験をSTAP細胞からダイレクトにできたなんて書いて通るはずがない。小保方さんを山梨に連れてくるために言った勧誘言辞をそのまま利用して、そんなこととはつゆ知らない無関係な笹井さんに押し付けたことになる。山梨に来てくれたらこの幹細胞化の論文は小保方酸浴細胞の核を使用したntES化実験だと本当の研究論文を書かせることができる。でも、それを知らないままに笹井さんが論文を書いたら、いくら何でも通るはずは無いと考えたんですね。それで強硬にネイチャーへの二報同時報告に拘った。例えばPNASに笹井さんの論文構成力と信用力で投稿したら通るでしょう。そしてキメラができたということに焦点が当たる。そこは自分の責任のあるところとなる。困るんですね。だから何が何だか分からなくする意味で幹細胞化論文の二報同時投稿を強硬に主張したんですね。
ここでどうして本当のことを言わなかったのでしょうかね。やっぱり人事のことでいろいろと自分で売り込んでるんでしょうね。特に西川さんがメンターですからね。今まで話してきたことと矛盾するようなことがあると話せないですね。論文が通らなければそれが一番無事なんですね。
小保方さんに幹細胞が自分ではできないと訴えられても、作り方を目の前で教えもしていない。論文は通ったら困るんですね。こんな論文はトンデモなんだから通るはずはない。通るように教えるなんて輪をかけてトンデモだ。僕の手技だよとごまかしてシカトしている。そんな科学は普通ではあり得ません。
若山さんの誤算は自分はntES化させたことを知っていて、ntES化させないでキメラができるわけが無いと分かっている。査読者たちは誰も信じないと思い込んだことです。現に三誌でリジェクトされている。でも、笹井さんはそんなことは知らない。ここで起きていることはすべてダイレクトに小保方細胞が引き起こしている現象だと信じて論文を構成している。
若山さんが通るなよ、通るなよと念じているのに、笹井さんは通るように通るように努力した。理研と笹井さんと丹羽さんの信用力が加わって、理研と若山さんだけの信用では通らなかった論文が通りそうになってしまった。
小保方ユニットリーダーの元には寺下さんがついていて、若山研とは常時連絡を取り合っている。若山さんは追い詰められてしまったんですね。
(出るべきバンド)幹細胞とキメラ体細胞からはどういうTCR再構成バンドパターンが出るべきなのか。CD45と酸浴STAP細胞からは全部のパターンが出る。こちらは簡単で、現に出ていますね。T細胞を選別しているのですから当たり前です。
でも、幹細胞とキメラは違いますね。まず幹細胞の方は、小保方さんの酸浴STAP細胞を基本ES培地で培養したら増殖を始めたということが本当なら、これはSTAP細胞と同じく全バンドが出ます。すべての幹細胞株でTCR再構成の全バンドがでる。それは細胞集団がポリクローナルな集団だからです。
ところが、この実験を若山さんは小保方さんにさせなかった。これを確認したのは別のラボメンバーで、結果は調べられた8株にのうち2株にはTCR再構成があると、小保方さんは知らされているように手記の149Pに書いている。ラボの中での報告でも、それを合わせて報告していると思われるが、桃子本ではあったと本人が報告したと書かれ、桂報告書でも相変わらずあたかも小保方さんが全部行ったかの如くミスリーディングな書き方をしている。
8株のうちの2株にあるという意味はとても不可解なものです。論文のプロトコル通りに作られているものなら、すべての株に全バンドが出ないとおかしい。
一個一個の細胞のTCR再構成のありかたの可能性の全ては以下ですね。我々のお勉強の成果でした。もう一度確認しておきましょう。小保方さんのプライマーで挟むとカッコ内の6本のバンドが出る。7~12は検出できないということです。J1、J2ともセグメント数を6としています。実際には7つのようですが、再構成するときの仕組みが素人には理解がちょっと難しい。でもそれはこの問題にはあまり関係しない。
>>
01.D1-J1の123456-D2-J2の123456(*GL)
02.D1-J1の23456-D2-J2の123456(*GL)
03.D1-J1の3456-D2-J2の123456(*GL)
04.D1-J1の456-D2-J2の123456(*GL)
05.D1-J1の56-D2-J2の123456(*GL)
06.D1-J1の6-D2-J2の123456(*GL)
07.D1-J2の123456
08.D1-J2の23456
09.D1-J2の3456
10.D1-J2の456
11.D1-J2の56
12.D1-J2の6
13.D1-J1の123456-D2-J2の234567(*リアレンジバンド)
14.D1-J1の123456-D2-J2の3456(*リアレンジバンド)
15.D1-J1の123456-D2-J2の456(*リアレンジバンド)
16.D1-J1の123456-D2-J2の56(*リアレンジバンド)
17.D1-J1の123456-D2-J2の6(*リアレンジバンド)
ポリクローナルな細胞集団をPCRの検体にすると全部のバンドが出るということです。論文通りの幹細胞の作り方だとそうならないとおかしい。8株の中に2株ほど"あったようだ"というような出方にはならない。
しかし、検体がモノクローナルな細胞集団であると、この中の二つの組み合わせになる。というのは染色体が二本あるからです。従って、上記のリストの二つの組み合わせになる。結果的に0本か、1本か、2本ということです。
でも、小保方さんも、手伝ったラボメンバーも8株の検体がモノクローナルな細胞集団だなんて予期していませんよね。丹羽さんは後にこれをもう一度検査させて、おかしいと気付き論文から外すように笹井さんにアドヴァイズしました。流石ですよね。ただし、ntES化されているなんて考えもしてませんね。何か予期しない実験上の手違いがあると考えて後でゆっくりやり直したらいいと考えている。捏造なんて疑ったら手伝い自体をつづけられませんね。
キメラの場合はもっと複雑ですね。今度は小保方細胞そのものであっても、小保方細胞核使用ntESであっても、20個程度移植した個々の細胞の増殖していった器官部分は0本か、1本か、2本のいずれかのバンドしか出ない。狭い範囲を試料採取するとそうなるが、広い範囲で採取するとポリクローナルな結果と同様になる可能性もある。キメラの場合の尻尾の細胞がモノクローナルに構成されているのか、ポリクローナルに構成されているのか、事前に誰か研究でもしているのか。まして、2Nの場合はリシピエントの細胞も考えないといけない上に、組織の採取時点で、リシピエントからの白血球が混じってないという可能性も排除されているかどうかが書かれていない。方法論自体が厳密さを欠いている。丹羽さんと笹井さんが幹細胞とキメラのTCR再構成結果を外させたのは当然ですね。ただ、キメラはスタンダードなやり方でできているということは疑ってないから、捏造に気づくなんてことは誰にもできません。キメラは出来たと聞いていて、それが前提で論文の書き方を指導してやってくれと頼まれているだけだ。捏造だと思ったら途中で降りるでしょ。そして捏造調査に入るでしょ。
11jigen氏が笹井さんの立場に居たら、同時に国会図書館に行き、博論を調べ、ティシュー論文も調べて突合せるということを、論文のリヴァイズを行う傍らで3か月間で両方やれたというのでしょうか。そんなことを疑うのだったらこの仕事は引き受けないでしょう。そもそも11jigen氏が博論を国会図書館にまで調べに行って、コピーまで取ってきたのは自分の自発的行動なのか、それとも誰かに情報を齎されてのことでしょうか。
(オスしか居ないことを不思議に思わない不思議)2NキメラのTCR再構成PCR結果がGel2と特許申請書に存在している。これがとても錯綜した結果です。Gel2の2Nキメラです。一本欠如がある。難問です。
取り敢えず幹細胞の雌雄の問題から検討してみましょう。桂報告書の検査結果は以下です。
>>
①FLS1~8 オス
②CTS-1,11~1 オス
③GLS-1~13 メス
④AC129-1,2 オス
⑤FLS-T1,T2 オス
⑥GOF-ES メス
⑦129B6F1ES1 オス
⑧129/GFP ES オス
⑨FES1 オス
⑩FES2 オス
⑪ntESG1 オス
⑫ntESG2 オス
まず、FLSは現状は全部オスです。Article Extended Data Figure 8-jがそのFLS8ラインの4Nキメラのジャームライントランスミッション実験の結果です。
以下がそのリジェンドです。
>>
j, Production of mouse chimaeras from STAP stem-cell lines by the tetraploid complementation method. *These STAP stem-cell lines were generated from independent STAP cell clusters. 4Nキメラということは胎児の体の組織すべてがオスだということです。従って精原細胞もオスです。要するにオスのマウスということです。
8ライン由来4Nキメラは全部雄です。従って兄妹交配は出来ません。相手のメスにGFPの無いものを持ってこないとジャームライントランスミッション確認ができない。「僕のマウス」のGFPがどうこう言う以前に、この時点ですでに若山さんがどうしてオスだけなのかということを不思議に思わないこと自体が彼の嘘を証明しているんです。8ライン作って全部オスなんて、仮に正しく小保方さんのスフィア細胞を使って作っていても、或は我々が判断しているように、小保方さんの細胞の核を使ってntES化したにしても全部がオスだったなんて確率はほとんどありません。そうではなくて、一つの成功したntESがたまたまオスでそれを8つに株分けしただけか、クローン胚はたくさん作っているはずだから、成功しているntESラインは複数あってそれを持っているのだが、小保方さんにこれがFLSの8ラインだから無限継代が可能かどうかちゃんと確認しなさいと渡していた8ラインが成功した複数のntES株の中の一つだけを8株に分割しただけのものであるかのどちらかでしょう。
既に、桂報告書が調べたFLSが何であるかは明らかになっています。小保方さんが普段使っていたFLSです。


松崎氏が2014/10/21に持ち出して調べたFLSサンプルは木星リスト123番なんです。13というのは上のGLSの13です。小保方さんが間違えてこちらに入れてるんです。FLSは8ライン、GLSは13ラインです。このFLSの全ラインがオスだったということです。でも、ここで雌雄を調べたとは限りませんね。なぜならこの調査は若山さんが先回りして無断で行っていますよね。以下です。

FLSが8株全部オスだということは調べなくても若山さんは知っています。なぜならFLSキメラにメスがいなかったから129/SvのGFP無しの雌マウスと戻し交配してジャームライントランスミッション実験をしたんです。「STAP細胞は産仔を数匹混ぜて作るため、性別が一致する率は高くない」って、今更何言ってんでしょうか。ジャームラインの実験の時には分かってやってたんでしょうよ。
そして、この頃に生まれた子の半分にしかGFPが来なかったと小保方さんに言った。つまり、この実験が何かコンタミがあったかもしれないという言い訳をこの時に準備しているんです。それはこの段階ではもっぱら米国特許仮申請をしてしまったヴァカンティに対する言い訳です。
又、小保方さんがリヴァイズ中に丹羽さんとPCR再検証してTCR再構成が無かったとされた実験後のサンプルが、木星リストの2番、4~10番のP2とされている8ライン分ではないでしょうか。笹井さん丹羽さんはこのサンプルを世界に公開しようとしてプロトコルを書いたんですね。その時にこの幹細胞にはTCR再構成は無いと書いた。手記の説明はそうなってますね。キメラ実験を信じているんですから当然の処置ですね。
それをTCRがないのに捏造を疑わないとは何事だと騒いだ最初が遠藤氏でしたね。舐めてると書いてましたね。無関係な人が突然入り込んできて丹羽さんに対して舐めてるなんて学者とも思えないやくざめいた唐突な書き込みが、kahoの日記でした。この人が何者なのかは11jigen氏と同様いずれ検討されなければなりません。
ともあれ、我々のntES仮説であると一種類だけを株分けすると全株がオスだけということは当然あるのですが、先に述べた2Nキメラの尾部のTCRに欠如しているバンドがあって、これは何種類かのntESラインを混ぜている可能性しか考えられないところなんですね。リシピエントの血液が混じっていたら全バンド出ます。
キメラのバンド欠如を考慮すると、我々はたくさん作ったntESをいくつか混ぜてキメラ胚に挿入したのではないかと推測したのですが、
幹細胞に関しては1種類を8株に株分けしただけのような結果になっている。桂調査は太田ESのコンタミ捏造という線で論旨を構成しているのでntES説に関しての証拠が不足していて、なんとも分かりにくいところです。ただ、今我々は太田ESのコンタミは無いと証明してしまってますからね。ここは本物かntESしかありませんが、幹細胞がポリクローナルな集団で無かったことで既に、論文通りの幹細胞でないことも証明されてしまってますよね。幹細胞はモノクローナルな細胞株なんです。桂報告書27P。
>>
1)TCR 遺伝子再構成に関する不整合データ隠蔽の疑いについて
(調査結果) 小保方氏は TCR 遺伝子再構成に関する実験を開始し、STAP 細胞を含む細胞塊、一部の STAP 幹細胞に TCR 遺伝子の再構成が見られることを CDB 若山研で最初に報告した。しか し、後に 8 系統の STAP 幹細胞の TCR 遺伝子の再構成を確認したところ、再構成は確認 されなかった。なお、この8系統は小保方氏が継代培養を繰り返していた細胞であった。
さらに、この実験は小保方氏の依頼で、CDB 若山研メンバーによる TCR 遺伝子再構成 の確認実験が行なわれた。しかし、この CDB 若山研メンバーの実験ノートによれば、実 験の結果 TCR 遺伝子の再構成は確認されなかった。
以上のことから、小保方氏は最初の実験でTCR遺伝子再構成があることを報告したが、 後の小保方氏自身の実験、および CDB 若山研のメンバーに確認を依頼した実験では TCR 遺伝子の再構成を認めるに至らなかったことから、実験データに不整合が存在したこと は明らかである。
丹羽氏は 2013 年 1 月に論文作成に加わった際に、小保方氏が継代培養を繰り返して いた 8 系統の STAP 幹細胞の TCR 遺伝子の再構成は確認されなかったと聞いたと説明し ている。さらに、丹羽氏は笹井氏に対して、TCR 遺伝子再構成に関するデータを論文に 含めることについては慎重にすべきとの意見を伝えた。小保方氏の追試が不成功であっ た点に関して、笹井氏らは STAP 幹細胞がヘテロな集団であり、長期的な継代培養により再構成が起っていた細胞が消失したという解釈を採った。なお、Article 論文には、 STAP 細胞を含む細胞塊の TCR 遺伝子再構成については記載されたが、STAP 幹細胞自体 の TCR 遺伝子再構成実験の結果については記載されなかった。
一方、丹羽氏は、Protocol Exchange への投稿は、発表後、この論文ではすぐに再現 性についてクレームがつくと思った。小保方氏のプロトコールでは不十分と考えそれを 詳細にしたものを早急に公表すべきと考えた、と説明した。さらに、当時、小保方氏と 笹井氏はコリジェンダム(corrigendum)で相当に多忙であり、エディターと応答でき る者が必要ということで、自分が執筆した、と説明した。
2014 年 3 月 5 日に Protocol Exchange に公表された詳細なプロトコールの「STAP stem-cell conversion culture」「 2.After 4-7 days of…」のプロトコールの「IMPORTANT」 (iii)に、8 系統の STAP 幹細胞には TCR 遺伝子再構成が認められない、という結果の記 載が存在している。
また、丹羽氏は「若山さんは、最初 STAP 幹細胞の初期のパッセージでは TCR 遺伝子 再構成はあった、と小保方さんから聞いたと言っている」と説明した。
(評価)
TCR 遺伝子再構成に関しては、最初小保方氏が再構成を確認したとされたが、その後 の CDB 若山研メンバー、および小保方氏自身の追試で失敗した。その事実にもかかわら ず、実験結果を自分たちのアイデアに沿うようなものを採用したものの、後に、Protocol Exchange で 8 系統の STAP 幹細胞には TCR 遺伝子再構成が認められないという結果が記 載されたこと、並びに丹羽氏への聞き取り調査における上記の説明から、意図的な隠蔽 ではなく、研究不正とは認められない。 小保方さんの手記での説明と矛盾していますね。手記で書いている幹細胞サンプルを世界に公開するという目的があって、プロトコルにはTCR再構成が無いと書いたのだという事情を桂報告書は意図的に外していますね。そしてTCR再構成が無いということを捏造だと気付くべきことだと世間に間違った印象を与えるように文章を構成している。キメラが出来ているという大前提の下で幹細胞のTCR再構成実験に何らかの間違いがあることは、後にちゃんとやり直せばいいことで、キメラの捏造を疑うのならまず若山さんが何をしたかから調査しなければならない。しかし、彼らは若山さんの主張している通りに調査を始めた。犯人の指示に従って調査しているんですね。御笑い種です。警察を入れないからこんなことになる。
「さらに、この実験は小保方氏の依頼で、CDB 若山研メンバーによる TCR 遺伝子再構成 の確認実験が行なわれた。しかし、この CDB 若山研メンバーの実験ノートによれば、実 験の結果 TCR 遺伝子の再構成は確認されなかった。 」と書きながら、だから幹細胞にTCR再構成があるなんて書かれてないでしょと言うことに気づかないわけも無いと思われるが、もしここで、桂チームが丹羽さんたちが気づくべきだと思ったのなら、どうして若山さんが何をしたかを調べなかったのか。何を勝手に太田ESのコンタミだという若山さんの嘘に騙されているのか。若山さんが犯人だということに気づけば調査サンプルの中身が入れ替えられているということにも意識が向くでしょう。笹井さんたちがこの実験が捏造だと気付かなかったことをとやかく言う前に、お前たちがなぜ若山さんが犯人だということに気づかなかったかを先に反省すべきだと、警察なら言うでしょうね。プロは人を疑う時には先にすべてを公正な立場で検証する。第三者調査といいながら、若山さんが犯人だという可能性を最初から除外している。捜査のドシロウトと笑われてしかるべきでしょうね。尤もど素人だったからなのか、意図的な隠蔽だったのかはこれから暴かれていくでしょうね。
今、Ooboe さんのパートナー氏がその細胞の入れ替え問題に関して検察に申請書を提出された。ES細胞を小保方さんが若山さんに気づかれずに渡すことはできないと既に証明されている。にも拘わらずFES1はFLSと同じだという調査結果がでた。ブーメランですね。これがサンプル入れ替えの動かぬ証拠です。FES1の中身は若山さんが入れたFLSです。このことは和モガさんが既に近縁率の数値結果からも導いていますね。和モガブログで確認されてください。(細胞増殖率グラフ)FLSに関してついでですから調査されていないP40のことに触れて置きましょう。FLS1~8の細胞増殖率表は小保方さんが正しく3日ごとに40継代しているからP40のサンプルが残されているんです。もう一度掲載しましょう。

小保方さんはこの中の一つのデータを増殖率表に使用したんです。桂報告書が海外出張等で3日に一度出られた日はないと言ってるのは小保方さんがES細胞の増殖率データに関して、2011年の春と嘘をついたから、まだ腰かけているだけで客員にもなってないときですから当たり前ですが謝金支払表になかったんです。
ES細胞の増殖率なんてわかり切っていてわざわざ金をかけて新たにやらなくてもデータはどこにでもある。若山研の予算でそんな無駄なことを若山さんがやらせるわけがない。小保方さんは咄嗟のことで事情も分からないままに、若山さんを庇うための嘘をついた。桂報告はそれを承知の上で捏造指摘している。大事なのはSTAP細胞の増殖率とFLSの増殖率で、それはちゃんと行われていて、Ooboe さんのパートナー氏の取り寄せた謝金支払表ではほとんど出勤している。ついでですから証拠保全をあちこちに置くという意味でも楠本さんのデータをここにも貼り付けて置きましょう。
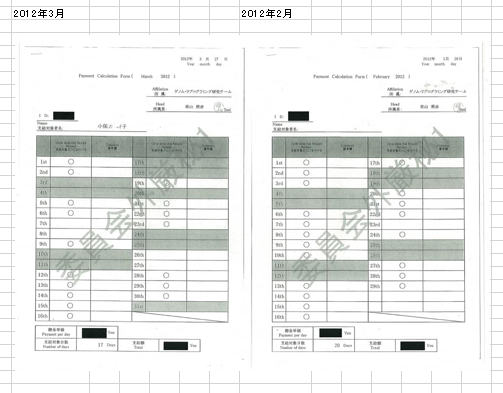

桂報告書はこの件に関して以下のように結論している。ESの増殖率実験が実際には行われていないことを、FLSが行われているにも関わらず意図的にレトリックを弄してあたかもどちらも行われていないかのごとくに書いている。
>>
(評価) この実験は行われた記録がなく、同氏の勤務の記録と照合して、Article Fig.5c のよ うに約 3 日ごとに測定が行われたとは認められない。"この実験"というのは正確にはES細胞の実験のことだけを言ってるんです。でも勤怠を見てない人はFLSも行なわれていないと思うでしょう。でも上のサンプルに小保方さんの字でP40と書かれたFLS-1~8があります。そして2月から5月にかけての120日間3日ごとに出勤できないような箇所はありません。そもそも腰かけていた2011年の春はまだ蛍光細胞すらできてなくて、こんな時期に分かり切ったESの増殖率実験を小保方さんがやってるわけがないし、まだ客員にもなってないのに勤怠があるわけもない。そんなことは桂チームも分かってるんです。小保方さんが何が起きたのか分からない状態で、とっさの嘘の言い訳をついたのを利用して間抜けなレトリックを弄して捏造虚偽報告しているのです。
(再度雌雄の問題)本題に戻って、今ある通りにFLSが全部オスだけだったらメスは生まれてこないんで兄妹交配はできないから別のジャームライン確認用の雌を持ってこないといけないと分かったところです。Article Extended Data Figure 8-j は4Nの実験だからそうなる。では2Nだったらどうなるのか。それがひとつ前の図のArticle Extended Data Figure 8-i です。
リジェンドは以下です。
>>
i, Production of chimaeric mice from STAP stem-cell lines using diploid embryos. *These STAP stem-cell lines were generated from independent STAP cell clusters.ここにはメスがあります。なぜならリシピエント卵のインナーセルマスから雌ができるからです。GFPがあろうとなかろうと兄妹交配してGFPが来たらジャームライントランスミッション有、つまりYesとなる。
アーティクル本文のマテメソに以下のようにある。
>>
Because the number of CD45+ cells from a neonatal spleen was small, we mixed spleen cells from male and female mice for STAP cell conversion. To make germline transmission more efficient, we intercrossed chimaeras in some experiments.ここで小保方さんが書いていて、かつ笹井さんが英文をみてやったintercrossの意味は兄妹交配(sibling mating)のことで、backclosss(戻し交配)と対照的に使っているのではないかと思われますが、正しい使い方か否かは素人なのでよくわかりません。近交系マウスを作るときにはincross,cross,backcross,intercrossという概念があって、それぞれ、雌雄相互に同一遺伝子をホモにもつ個体同士の掛け合わせ(完成した近交系マウスの掛け合わせはすべての遺伝子座でインクロスになる)、雌雄相互に異なった遺伝子をホモに持つ個体同士の掛け合わせ、ホモとヘテロの掛け合わせ、ヘテロ同士の掛け合わせという概念です。
この定義に従うと、STAP細胞の場合、そもそもF1マウスを使っている段階から先はできた子供同士の掛け合わせやワイルドタイプとの掛け合わせは全部インタークロスになります。そして別の近交系マウスを持ってきて掛け合わせたときだけバッククロスしていることになる。用語として正しいのかどうかはわかりませんが、他方メイティングの実際は論文で出ているわけです。
以下はArticle Extended Data Figure 7-bと7-cのSTAPキメラとそのジャームライントランスミッション実験の表です。

リジェンドは以下です。
>>
b, Generation of chimaeric mice from STAP cells by cluster injection. STAP cells used in the experiments above were generated from CD45+ lymphocytes of multiple neonatal spleens (male and female tissues were mixed). *All fetuses were collected at 13.5 d.p.c. to 15.5 d.p.c. and the contribution rate of STAP cells into each organ was examined by FACS. **The contribution of STAP cells into each chimaera was scored as high (>50% of the coat colour of GFP expression). ***B6GFP: C57BL/6 mouse carrying cag-gfp.
c, Production of offspring from STAP cells via germline transmission. Chimaeras generated with 129/Sv × B6GFP STAP cells (obtained from the experiments shown in b) were used for germline transmission study. b図は2012年初頭の実験です。DBA/2とのF1も使われている。この実験は2Nキメラ実験です。リジェンドのaにも書かれているんですが、右端にドナー貢献度が書かれているからそれだけでも判断できます。4Nキメラであれば胎児は全部ドナーですからこの記載は不要です。
そしてこのb図の2段目の129/Sv x B6GFPのキメラのジャームライントランスミッション実験がc図です。b図でキメラになっているのが20匹(内 High contributionが6匹)です。
2Nキメラですので何を根拠にキメラになっていることを調べたかというと、注に**The contribution of STAP cells into each chimaera was scored as high (>50% of the coat colour of GFP expression).とあって、GFP蛍光している毛色の面積が50%超をHigh contributionとしているので、その他も体毛のGFP蛍光判断です。そして紛らわしいですが、*All fetuses were collected at 13.5 d.p.c. to 15.5 d.p.c. and the contribution rate of STAP cells into each organ was examined by FACS.と書かれているのはあくまでも、最上段のB6GFP x DBA/2の子供だけで、これは帝王切開で全部取り出して、各器官組織を取り出して解剖し、FACSでGFP貢献度の確認をしたということです。従って二段目の129/Sv x B6GFPは出産されたマウスで、GFP蛍光の確認は体毛だけです。
この20匹から9匹を選び、ワイルドタイプを1匹加えてメイティングさせた。それがc図です。
2Nキメラですから体毛が蛍光しているからと言って生殖細胞までドナーだとは限りません。体毛判断でHigh,Medium,Low と分けているんですが、何か意味があるのかどうか分かりません。精原細胞、卵原細胞というのはとても早い段階で偶然の位置関係で決まるのではないですかね。毛色の貢献度とどういう関係なのか、よくわかりません。
結果、105匹の子供が生れて体毛のGFPもしくは黒目判断(リシピエントのICRはアルビノなので赤目)でジャームライントランスミッションの確認されたものが35匹ということです。これは見えるところにありさえすればいいので、見えないところで(例えば心臓で)ひょっとしたらあるかもしれないですが、実験全体としては一つでもあればトランスミッション有ということなので他は関係ない。三段目の実験は32匹出産されて体毛と赤目判断では0でしたが、それは心臓にはGFP蛍光があったかもしれないが、他で既にトランスミッション確認されてますからSTAP細胞はキメラ形成能だけでなく、更にジャームライントランスミッション能もあるという証明終わりだということです。
幹細胞キメラは別ですが、このSTAPキメラに関しては、すでにTCR再構成のPCR結果がサイエンスで書かれていることは査読文書で分かります。
>>
The DNA analysis of the chimeric mice is the only piece of data that does not fit with the contamination theory. But the DNA fragments in the chimeras don’t look the same as those in the lymphocytes. This assay is not properly explained. アーティクル論文とは違っているようですが、キメラのTCR結果も入っていますね。これらを丹羽さんと笹井さんが外させたんですね。
(B6の3番染色体上の5bp欠失とキメラ子8のGFP欠失)さて、問題はこの時のキメラ子9匹のDNAが残されていることです。桂報告書10Pです。
>>
(調査結果) 1)Article Fig.4 と Extended Data Fig.7 に 129/Sv×B6(CAG-GFP) F1 マウスから作ら れた STAP 細胞由来の 2N キメラができたこと、さらに germline transmission により、 このキメラの子ができたことが報告されている。 小保方研のフリーザーに「カルスキメラ子 1」~「カルスキメラ子 9」と書かれた 9 本の DNA 試料があり、2011~2012 年の CDB 若山研では STAP 細胞を「カルス」と呼んで いたことから、これらはこのキメラの子の DNA と考えられた。実際に、小保方氏への聞 き取り調査により、これらの試料は Article Extended Data Fig.7 に出てくるキメラの 子から小保方氏が抽出した DNA であることを確認した。若山氏の実験ノートでは、この キメラの作製は 2012 年 1 月終りから 2 月はじめにかけて行なわれていた。
9匹なんです。ジャームライントランスミッション実験時に親として選ばれたキメラがWild type(Wide typeはタイプミス)を除いて9匹でした。でもこれはキメラのF0です。キメラ子という以上F1でしょうね。つまり35匹生まれた中の9匹でしょう。この検証結果が桂報告書のスライドにある。こちらはBCA報告にはないものです。以下です。
まず、3番染色体にAcr-CAGの入っていたカルスキメラ子は2,3,6です。このGFPが入っているのは岡部マウスです。岡部マウスというのはB6マウスです。F1マウスのB6側の3番染色体にこのAcr-CAGが入っている。同じく3番染色体には5bpの欠失があってそれはカルスキメラ子2、3、6、8にあるのですが、実はこれもB6側の3番染色体なんです。同じく桂報告書のスライドです。
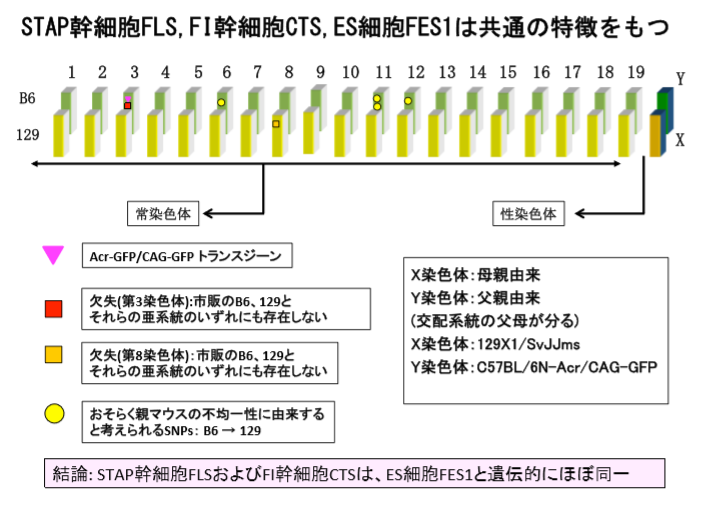
ピンクの下向き三角と赤の四角です。
B6側のGFPと欠失は同じ3番染色体上にあるので一緒に動きます。カルスキメラ子の8にはGFPがついてきていません。不思議です。
原因として通常考えられるのは減数分裂時の乗り換えでしょうか。2Nキメラのジャームライントランスミッション時に、できた子供の一つに、GFPが減数分裂時に相同組み換えを起こして別の染色体上に交叉融合して飛んでしまっていたものが居たということになる。しかしGFPはこの場合PCRで探しています。岡部マウスと分かってしまっているのですから既知のプライマーで挟んでます。別の染色体に飛んでいても存在していたらPCRにかかってくるはずですね。乗り換えではなさそうです。
因みに5bp欠失の見つけ方としては、場所はシーケンシングで分かっていますから、その場所の前後近辺のプライマーを作ってPCRにかけて挟むと長いのと短いのが出るので短い方が4つバンドになっているところが提示されているわけです。
GFPに関してはマウスコロニーにGFPの無いB6が飛び込んだのか、減数分裂過程、もしくは培養時の体細胞分裂過程でGFPの欠失が生じた等の可能性が考えられるが、5bp欠失は4つに共通していて1つだけにGFPが無いという現象がどの可能性を許すかという問題になる。GFPの有るマウスと無いマウスのどちらにも5bpの同じ欠失が生じるということは無いので、やはりGFPのあった細胞に5bp欠失が生じ、それをいくつか株分けして継代培養しているうちにその一つにGFPの欠失が生じたと考えるしかない。できたばかりのSTAP細胞を移植してこういうことは起きない。我々のntES仮説を支持している数々の証拠の中の一つの現象ではないか。
因みに太田ESは作っただけで使いもせずにすぐ凍結して、解凍するまでは時間は止まっていたので、それに培養変異があるなんてことはない。桂報告の小保方さんによる太田ES使用捏造結論でも、変異が入るなら小保方さんが解凍した後の話ですね。若山さんのntES化であっても、小保方さんの太田ES使用であっても、変異期間は同じ長さです。桂報告書の論理はとても非合理なもので科学者の書いた文章とは思えないところです。小保方さんは光っているマウスだけを9つ選んだと推定したが、PCRの結果は光っているのは3つしか選んでない。キメラ子自体は105匹生まれていてその中の毛色でジャームライン有になったもの35匹の中から9つ選んでいると考えたのですが、このPCR結果は105匹の中からランダムに9匹選んだことになる。それで35/105はほぼ3/9で計算上はあっています。
これらカルスキメラ子105匹のマウス背景は何であり得るかというと、元の2Nキメラの生殖細胞はドナーである129B6であるか、リシピエントであるICRであるかのどちらかです。毛色でキメラになっているもの同士の兄妹交配ですが、GFP蛍光する生殖細胞以外はどうであるかは実際には関係ありません。
配偶子は129とB6そしてICRとICRに減数分裂する。たまたまICR同士がメイティングしていたら子供はすべてICR/ICRが生まれる。129B6同士だったら129B6が2、129/129が1、B6/B6が1の比率で生まれます。ICRと129B6がメイティングしていたらICR/129が2とICR/B6が2生まれます。B6の入っているものだけが光る。
因みにキメラの子はキメラではありません。普通のマウスです。
岡部マウスは太田さんが岡部さんの元から若山研に来た2003年からあるはずのものです。彼は精子での研究に来ている。無論自分の研究のために若山研に持ち込んだものなので、それ以来若山研で維持されている。
「僕のマウス」の親の129ではとんでもないマウスコンタミがありました。岡部マウスに関してもコンタミ懸念は無いのか。マウス会社から別のB6を買ったことがあるのは記者会見で証言もされている。
ただ、手記によると若山さんは光る精子を集めていたと書かれているので、岡部マウスの交配は顕微授精で行われていた可能性もある。卵はCAGで光るし、精子はCAGで光らない代わりにアクロシンで光る。このやり方で継代し続けている限り、比較的マウスのコンタミは排除される。特に他の遺伝子コンタミは防ぎえないかもしれないが、GFPに関してはこれの無いB6は取り除かれてしまうのでGFPのないマウスは無いことになる。
桂チームは2010年に若山研で凍結された岡部マウスの受精卵を調べて3番の欠失は無いと言っています。どうして最新維持されている岡部マウスは調べなかったのか。小保方さんの研究のための実験は2011年から2012年にかけて行われた。2010年の凍結卵だけ調べては分かりません。こんなついでに調べられるような今の岡部マウスを調べないというのは結論ありきの線から外れそうなものはもともと調べないという方針だと疑われても仕方無いでしょう。
桂報告はFLSは太田ESのFES1だということにしてしまったのですが、FES1は2005年の論文で使われた129と岡部マウスを掛け合わせた129B6F1のntES-G1G2、もしくはその元を作った頃に、何の目的もなくただ同じマウスから受精卵ESを作ったと、本人によって物語られているもので、歴史的な証拠は何もないものです。本人は日経サイエンスの取材で細胞リストに書かれていると証言しているようですが、その開示はありません。因みに理研に提出されているntES-G1G2のラベルには129B6F1G1と129B6F1G2と書かれているのですが、理研の調査結果ではどちらもB6/129で親の雌雄が逆になっていました。太田論文には129/B6となっています。つまり論文記載背景とラベルは一致しているが中身が違っているわけです。
G1,G2と名前を付けたのが2008年かどうかは調べられていないから分かりませんが合理的推測では2008年です。先に作られたものに最初から2番と書かれているわけはない。中身が論文ともラベルとも違っていると分かっていながら証拠の信憑性確認をスルーするというのは詐欺犯罪に近い行為ではないでしょうか。
(培養変異)繰り返しますが、太田ESが2005年の論文のための実験中に作られていて2014年に調べられるまで10年近いから培養変異が入りやすいという理屈は無論嘘です。凍結されている間細胞の時間は止まっています。変異は培養回数によるんです。何日間培養され何回植え継がれたのかということはどのサンプルも調べられていません。そもそも実験ノートの提出公開は一部といえども小保方さん以外は行っていません。小保方さんの提出した実験ノートを公務員法違反行為と知りながらNHKに全コピー流出させた松崎GDを起訴もしないという、これも同時にそれ自体も公務員法違反である行為を犯していながら、小保方さんの実験ノートが提出されなかったから分からないなどと、とぼけたことを書いている桂報告を放置している理研は、追々追及を受けることになるでしょう。
FES1,2は2005/12/7に凍結されている。何の目的もなく作ったと太田さんが証言している以上、できたのち長く継代培養することはない。作成したのは12月中でしょう。ntESG2の凍結は2005/1/20です。論文を提出して既に2005/4/26にはアクセプトされている。その後8か月も経過してから何の目的もなくFES1,2を作ったと言ってる。余った129/Sv-terを使って実験したのかと思いきや、ここではX1を使った。若山さんが別途購入していたものを流用したのでしょうかね。ただ、その場合は2005年の論文とは何の関係もありませんね。何のためにntESG1,G2を提出したのか。報告書4P。
>>
また、STAP 幹細胞 FLS が、Acrosin プロモーター下に GFP を発現する Acr-GFP と、CAG プロモーター下に GFP を発現する CAG-GFP の共挿入を含むことが判明した後に、 過去に CDB ゲノム・リプログラミング研究チーム(以下「CDB 若山研」という)で作製さ れた Acr-GFP/CAG-GFP 共挿入 ES 細胞を取り寄せて解析したのが、下段の 4 種類の細胞で ある。アクロシンが出たから過去に岡部マウスを使った論文関係の細胞を出せと言われて太田氏と若山さんが理研に出したのが、FES1,2とntESG1,G2という書きざまですが、Ooboe さん提供データで明らかになっているように、これは桂調査チームより先に若山さんから東北大に解析依頼提出されている。むちゃくちゃですね。ガヴァナンスも何もあったものじゃない。
FES1は無かったはずのものです。報告書14P。
>>
ES 細胞混入のもう 1 つの謎は、ES 細胞 FES1 がどのようにして STAP 細胞研究時の CDB 若山研に存在したかである。ES 細胞 FES1 は 2005 年に当時の CDB 若山研メンバー によって樹立されたが、その後、研究に使わず、2010 年 3 月(CDB 若山研で STAP 研究 が始まる前)に転出した時に ES 細胞 FES1 の凍結保存試料を全部持ち出して CDB 若山 研には残さなかったとされている。当時の CDB 若山研メンバーへの質問状と聞き取り 調査、および関係者の実験ノートの調査でも、当該メンバー以外に ES 細胞 FES1 を使 用した者は見つからなかった。 だったらFES1は使われていないと分かりそうなものですがね。
FES1には3番と8番に欠失がありましたが、8番は129側ですので、今はB6の3番の欠失です。FES1の3番の欠失はいつできたか。2010年の岡部マウスの凍結受精卵にはなかった。無いのにどうしてそれ以前の2005/12/7に作られている太田ESにB6の欠失があるのか。それは培養時にできたのだという解釈ですね。培養は2004年の実験中と、2007年の解凍後の再使用時の培養の二度です。何回植え継がれたか調べられていない。一方、FLSやCTSは2012年の初頭から秋口までと笹井研での再解凍培養の二度です。こちらより太田ESの培養期間が長いとも思えませんね。どちらにも3番の欠失は起こり得るんです。そしてFES1と称してFLSを中身に入れ替えたらこんなことは当たり前の結果です。SNPs近似はむしろそれを指摘しています。これは和モガさんがずいぶん以前から指摘しているし、佐藤さんの本でも書かれている。
この3番の欠失は当然ですがテラトーマからも出ている。
>>
(f) STAP 細胞から作製されたテラトーマは、ES 細胞 FES1 に由来する可能性が高い
(調査結果) Article Fig.2eと Extended Data Fig.4a-c に登場する STAP 細胞由来のテラトーマは、 いずれも Oct4-GFP+細胞の 7 日目細胞塊から由来したとされている。しかし、以下1) ~3)の検証結果に示す通り、このテラトーマは、 (1)Acr-GFP 遺伝子を含むが Oct4-GFP 遺伝子は含まないこと (2)ES 細胞 FES1 に特異的な 2 個の欠失が定量 PCR の解析で検出されたこと (3)組織切片の FISH 並びに染色体ペインティングで大部分の細胞に X 染色体と Y 染 色体各 1 本が検出されたこと(ES 細胞 FES1 が XY(オス)であるという事実と合致) が判明した。よって、これらの図に登場する STAP 細胞由来のテラトーマは、ES 細胞 FES1 に由来する可能性が高い。太田ESを渡されて騙されたと言ってる若山さんの嘘は既に証明しています。ESを渡されたら大きさの違いで誰にでもわかる。太田ESでないのにアクロシが出るのは最初から小保方さんに渡しているマウスが129/Sv-X1と岡部マウスとのF1だからです。
そして若山さんの当時維持していた岡部マウスには3番の欠失は2010年まで無かった。
無論、2010年の凍結卵にないものは2008年以前には無かったはずというのは、マウスがちゃんと均一に近交系に維持されている場合の話です。不均一ですとたまたまGFPの欠失しているマウスが選ばれなかったというだけの可能性もある。どうも「僕のマウス」の片親である129CAGホモのB6特異的SNPs領域が広くてマウスのコンタミが疑われている。白毛のマウスの中に黒毛が飛び込んでるとしたら、黒毛の岡部マウスに黒毛のGFP無しマウスが飛び込んでわからなくなったいうことはあり得ないことではない。
こういうコロニーの汚染はランダムに数匹抜き取って調べないと少ない数では分かりませんが、そもそも岡部マウスのコロニー自体は調査すらされてない。自家繁殖なんてさせてる事自体がどうなのか。もっとも、若山ラボの研究目的にとって大きな障害ではなかったということはあり得ますね。そんな研究室に、マウスのコンタミなんてあってはいけない小保方さんの研究が参加してきてしまったという予期しない事情があったということも推定される。でも所詮マウス管理、なってないんですよね。
マウス自体にGFP欠失の有るものが混じったとしたらそれは、2010年以前と以後と両方あり得る。
しかし、まあ、時々顕微授精もしていたとして、結構長く自家繁殖もさせているから、GFPがヘテロに入っていたマウスがあったという蓋然性もそれほど高くはない。すると培養時にGFP欠失が起きたと考えることもできる。体細胞分裂時に起きた欠失ですね。
この場合は酸浴中、酸浴維持培養中の7日間、そして、この細胞の核を移植してntES化して後の培養中、更にその維持培養中に生じたと考えることになる。FLS樹立と同時に行われた2Nキメラでしょうね。テラトーマに最初のF1の幹細胞を注射してしまっていますから一からやり直したとすると日程的には以下のようになる。
2012/2/2***若山さんが129/B6-Act-CAGの赤ちゃんマウスを小保方さんに4,5匹渡す。
2012/2/8***小保方さんがSTAP塊を若山さんに渡す。
2012/2/9***若山さんがリシピエントの除核卵に小保方細胞核を入れたクローン胚をたくさん作る。
2012/2/13***数日後クローン胚が胚盤胞期になったところでインナーセルマスを取り出してntES培養を始める。成功率は1割程度。増殖成功しているシャーレはそれぞれ雌雄どちらかである。
2012/2/20***培養一週間後で増殖確認できたものをキメラ胚に入れる。いくつかのシャーレから数個ずつを混ぜて一つのキメラ胚に20個程度入れる。
2012/2/25***いくつかのキメラ胚のインナーセルマスを取り出してES培養し、幹細胞化する。これがFLSのライン。小保方さんに8ライン渡して増殖率実験を行わせた。120日後、2012/6/25前後に実験を終えて凍結されたものがP40のFLS1-8のサンプルである。
2012/3/10***20日後に2Nキメラの出産がある。若山氏実験ノートに2012/3/11とある。
2Nのカルスキメラ子8にだけGFP欠失がある。
ではこのGFPの欠失は何時起きたか。すでにテラトーマにはAcr-CAG-GFPが見出されている。しかし、テラトーマの場合はたくさん注入するので、作られた1ラインに欠失があっても他のにGFPがあれば検出される。問題は翌年の実験であるカルスキメラ子の8番だけにGFPの欠失が生じていることです。
我々はntES化されていると見ている。酸浴細胞をクローン胚に入れてntESしたいくつかのシャーレの中からランダムに取り出してキメラ胚に入れているとみている。そのシャーレのどれかでGFPが欠失したものができたと考えることはできる。その際はもう一つの欠失は岡部マウスに先にできていたと考えるしかない。2010年には無かったがその後できたと。顕微授精を常時行っていてもありうる。この場合今の岡部マウスを調べないと分からない。なぜ調べなかったか。若山さんが捨ててしまっていて2010年のものしかなかったか。
(不動の定点)これを考えるときに不動の定点があるのと無いのでは天と地ほどの違いがある。我々は今小保方さんによる既存ESのコンタミなんて無いのだと分かっている。ここが押さえられるまでは本当に悩まされたが今や、STAP cells are derived from ES cellsなんてトンデモだと分かっている。この表題だけならntESであることも含みうるが内容は太田ESと学生のGOF-ESと若山さんの「僕のマウス」ESを使った小保方さんによる捏造という結論になっている。大間違いですね。
もう一度貼り付けましょう。ニーチェはこういう時よく「驢馬どものための注」と書くのを好んでました。
左は最初の実験時に小保方さんの細胞をナイフでカットして胚盤胞に移植している写真です。若山さんが行ってる写真ですから、細胞塊は小保方さんが渡しているものです。左は通常の受精卵ES細胞です。大きさが違う。
少々胚盤胞の大きさが違っても影響しない。以下はマウスの胚盤胞期である3.5日目の解剖取り出しされた胚盤胞です。この後ハッチングと言って外側のたんぱく質の膜を脱ぎ捨ててエピブラストになってから子宮に着床しますから、前段階で外膜が柔らかくなって全体が1.5倍くらいに急激に拡大します。そして直後にハッチングする。このハッチング前の期間のExpanded Blastcystと呼ばれる胚をキメラ胚として使います。大きさが違うのは3.5日目で検討つけて取り出しますから、1匹に20個ほどある卵の段階が微妙に違うからです。一番小さいのは胚盤胞になり立てで、一番大きいのがハッチング直前です。この中から適当な大きさの径のパイプで胚盤胞を集める。こういうのはキメラ作成のプロトコルに説明されていますね。
実験には100個とか200個が使われますから一番数の多いサイズで集めて、集める過程で人の目で選択しているからやや小さいのも入りますね。入れる胚盤胞が少々小さくてもESの大きさと小保方さんの細胞の大きさの違いが分からないなんてことはありません。
3.5日の胚盤胞写真で一番大きいのと一番小さいのは直径で倍違いますね。一番大きいのと一番小さいのがたまたま選ばれている確率はとても低いですね。みな同じ作業していますから平均的に一番多い大きさを集めている。でも両極端比較もしておきましょう。
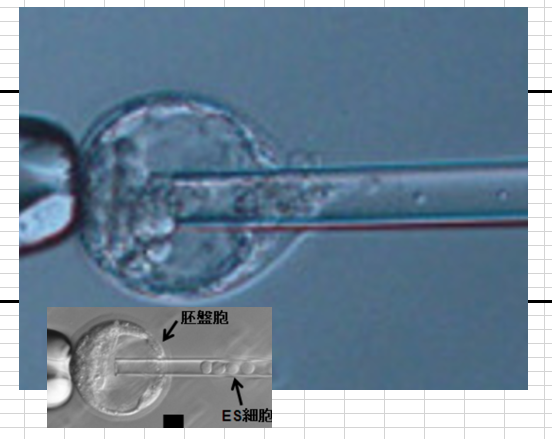
まだES細胞の方が大きいんですね。何年ESを見続けているのだということですね。小保方さんの細胞は基本リンパ球由来だから小さいんです。
直径で1/2、面積で1/4、体積で1/8です。同じ大きさにしようと思ったらESの写真はとてつもなく小さな胚盤胞を選んでいたということになりますね。以下のくらいかな。

こんな小さな胚盤胞を選ぶ人いないでしょ。そもそもSTAP細胞とES細胞の大きさは小保方さんが論文に写真を貼付している。彼女は形の比較を見せるために写真を同じサイズに調整していますから気づきにくいですが、左下にスケールがついているんですね。それを合わせたのが以下です。
ナイフ切り分け写真は後からのデモンストレーションとして撮影し直したかもしれませんが、彼は記者会見で質問に対してナイフ切り分けで入れた最初の実験でキメラができたんですと答えている。キメラの成功した時、彼はこんなに小さな細胞の幾分大きめの塊をナイフで切り出して、ES細胞の移植時より大きめのパイプを自分でバーナーで炙って引き延ばして、この細胞塊を移植したんです。そう自分で言ってるんです。もう一度貼り付けましょう。
失敗続きだったころ、彼はSTAP細胞の小さな粒に合わせたガラス管を作っていました。パイペットの作り方の図示は以下です。胚盤胞を左側で吸引して支持しているパイプをホウルディングバイペットといいます。カッティングニードルというのは若山さんが細胞塊を小さく刻んでいるあの針で、核移植の時にも使います。インジェクションパイペットが中にSTAP細胞やES細胞が入っているものです。


若山さんはSTAP細胞をトリプシンでばらして挿入している時はとても細いパイプを自分で作っていたことになりますね。彼のラボはntESキメラは常時作っています。受精卵ESもntESもその遺伝子解析的な性質は違いますが、大きさは同じです。普段ntES細胞は移植のためのパイプをいつも作っています。STAP細胞の時だけとても細く作る。細胞の大きさの違いを知らないということはありえませんね。
そして次に塊で入れたらその時に成功したから大きさについて意識していなかったといったんですがね。その後もその後も何回同じことしていることになるのでしょうかね。細胞の大きさ自体がかなり違いますよね。パイプとか胚盤胞との相対的な比較だけではありません。本当に何度もやっていたら大きさの違いに気づかないことはないでしょう。笹井さんがはっきり分かると言ってますね。
ナイフ切り分けなんてそもそも何度もはやってないと思いますね。最初の実験時だけですよ。
太田ESでないことが分かったら、ではなぜAcr-CAGがでるのか。それは若山さんが岡部マウスとのF1を使っていたからに決まってます。これはFLSを作った時、小保方さんに「僕のマウス」を渡したとという証言が虚偽だということを同時に証明しているんです。
手記208-209P。
>>
・・・若山先生は光る精子で実験をしていました・・・Article Extended Data Figure 7-bと7-cはSTAPキメラです。でも、若山さんの2011年11月までのキメラ実験失敗、検証再現実験で丹羽さんと小保方さんの作った酸浴細胞を清成さんがナイフ切り分けでキメラ胚移植実験した失敗結果からの我々の判断では、小保方酸浴細胞からキメラはできません。
従ってこのキメラはntESキメラだと考えるのが最も自然な推測です。
- 2019/09/05(木) 21:43:54|
- AC129
-
-
| コメント:0
(2) 疑念の検討A. コントロールES細胞a.AC129実験と多能性遺伝子解析実験の区別小保方さんはF1の受精卵ESは若山さんにもらった「僕のマウス」ESしか持っていません。それ以外に持っているのは学生のGOFのntESだけです。そして桂報告書によって太田ESを持っていたのではないかと疑われているわけです。しかし、ここでは太田ESは使われていないから関係ありません。
AC129というのは若山さんが129/Svの赤ちゃんマウスを小保方さんに渡して、STAP細胞を作ってもらって、それを培地誘導で幹細胞化させたと言ってるものです。129/Svを渡したのにAc129-1を遺伝子解析したら129B6F1CAG-GFP(ホモ)でしたという調査報告です。しかも、遺伝子欠失と重複の特徴からして、129B6F1CAG-GFP(ホモ)マウスから若山さんが作ったESの内のライン1だったということです。
それに対してSTAP ChIP lysateは、そもそも、今は雌雄表記の問題はわきに置いておいて、B6 x 129/SvのF1マウスだと小保方さんがデータベースに書いているのですから、129/Svの背景が出たらむしろおかしいわけです。GRASに提出して調べてもらっているのは多能性遺伝子の発現状況です。129/Svの実験とは基本無関係のはずなんです。
どうして調査チームはAC129の実験サンプルと多能性細胞遺伝子解析実験サンプルを混同したのか。それは竹市さんが言ったように最初にちゃんと調査対象のサンプルの出所と由来を調査しなかったからです。証拠の信憑性も含めた調査対象サンプルの証拠能力調査という、警察の捜査なら常識である手順を踏んでないからなんです。
サンプル調査は2014年の3月10日に若山さんが論文撤回を著者たちに申し入れたその日に、調査対象被疑者であるはずの本人が理研の許可もなく勝手に第三者と称する放医研の知人等にサンプルを送った。恐らく守秘を配慮して若山さんがMTAを結ばずサンプルを山梨大に持ち出さすことを積極的に依頼した手前、事件化して後は山梨大に依頼してサンプルをすべて差し押さえなければならない理研がMTAが無いことによって依頼できなかったという弱みをにぎられていたのである。さもなければ若山さんは試料を差し押さえられて勝手に解析に出すことはできなかったはずである。
その時に放医研に出されたのは以下です。
①FLS 8株
②FLS-T 2株
③AC129 2株
④GLS 2株
⑤コントロールES 5株
全て山梨大で彼が保有していた試料で、従ってAC129も若山さんの持ち出していた試料を送っている。この結果が6月16日の記者会見で発表されてGFPの挿入位置が18番染色体にホモに入っていて、コントロールESと同じだと分かった。調査は若山さんの主導で始まっていて、証拠能力の検証が先だと言った竹市さんと野依さんの意見は通らなかった。
AC129からF1背景が出たからAC129とF1が関係づけられて比較分析対象となったが、今残されているAC129が当時129/Svで行われたと言われている実験とどういう関係なのかということは調査されていない。ただ若山さんの証言を証拠もなく妄信しているだけである。ここに大きな疑念があるが、今は、細胞比較をリヴューすることから始めよう。
b.129マウスコロニーの不均一まずAC129が全解析されて、次に小保方研フリーザーにあった129B6F1 ES6が怪しまれたがために全解析を受けたと思われる。というのも仮に放医研から回ってきた129B6F1-1,2,3,4,6 ESの中の6だったら、これをまず全解析する理由はありません。佐藤貴彦氏の『STAP細胞 事件の真相』の中で、取り寄せた公開資料により、6月2日に以下の細胞が分析されていると書いています。又後のOoboeさんのパートナーさんの取り寄せ試料から管理室の持ち出し日も分かっています。
>>
①FLS-3(木星リスト5番 4/17 丹羽氏持ち出し)
②FLS-4(木星リスト6番 4/22 丹羽氏持ち出し)
③GLS-1(木星リスト28番 4/22 丹羽氏持ち出し)
④GLS-11(木星リスト38番 4/22 丹羽氏持ち出し)
⑤AC129-1(木星リスト24番 4/22 丹羽氏持ち出し)
⑥AC129-2(木星リスト25番 4/22 丹羽氏持ち出し)
①②はこのときに15番にGFPが入っていたとされたが放医研も理研も間違えた。GFPにアクロシンのプロモーターがついていたことからアクロシンの遺伝子は15番なので15番だと判断したが、これは岡部マウスのAcr-GFPの前につけられているアクロシンプロモーターで、実際には3番に入っていたことが後に分かったというお話になっているのはよく知られている。物事が順番に判明して行ったかの如くのお話になっている。ここで登場するヒーローもよく知られている人です。後程触れましょう。
因みに③④については丹羽さんが核型解析に出したんです。DORAさんの昔のブログにある。保存のためにこれもここに貼り付けて置きましょう。
佐藤氏によるとこの時にはすでに以下のサンプルも解析されていたという。
>>
⑦TSA ntES-9(木星リスト91番 6/9 松崎氏持ち出し)
⑧ES129 GFP+/-(木星リスト109番 6/9 松崎氏持ち出し)
⑨GFP ES(木星リスト131番 6/9 松崎氏持ち出し)
⑩129/GFP ES(木星リスト95番 6/9 松崎氏持ち出し)
⑪ES+/-(木星リスト109番 6/9 松崎氏持ち出し)
⑫129B6F1GFP ES6(木星リスト115番 6/9 松崎氏持ち出し)
⑫が全解析されたんですね。AC129は既に丹羽さんによって4/22に持ち出されている。最初丹羽さんは細胞の解析にも参加していた。しかし、後に外されて再現実験の担当になります。6/28に持ち出したのが最後で再現実験に移り、後を受けたのが松崎氏です。
因みに本日(2019/8/19)、Ooboe さんが学さんブログに以下のように書き込まれている。
>>
・・・6月2日にはStap関連と129B6F1ES6、129/GFP ESなどが解析終了してます。が記録にありません。この事実は重いのです。
竹市所長のしぶしぶ了承は6月5日です。竹市所長Goの前に持ち出しているのです・・・
2019/8/18(日) 午後 6:12[ Ooboe ]返信するこの件の経緯に関しては佐藤氏の『STAP細胞 事件の真相』(2016/12/14初版発行)の80P前後に既に書かれていることですが、Ooboeさんのパートナーさんは後にサンプルの調査持ち出し記録を入手されたのでその情報も付加されている。どちらも6月10日付の竹市センター長の報告書を根拠にされているが、
その内容が開示されていないので、サンプルが持ち出し記録にある6/9ではなくて6/2以前だということの確証がない。無論両方で主張されているので、その竹市報告に書かれているのだろうとは思いますが、早く開示されて欲しいものです。
参考のためにOoboeさんのパートナーさんが取り寄せた調査用のサンプル持ち出し記録を保存のために貼り付けて置きましょう。二列目が持ち出し者の欄です。
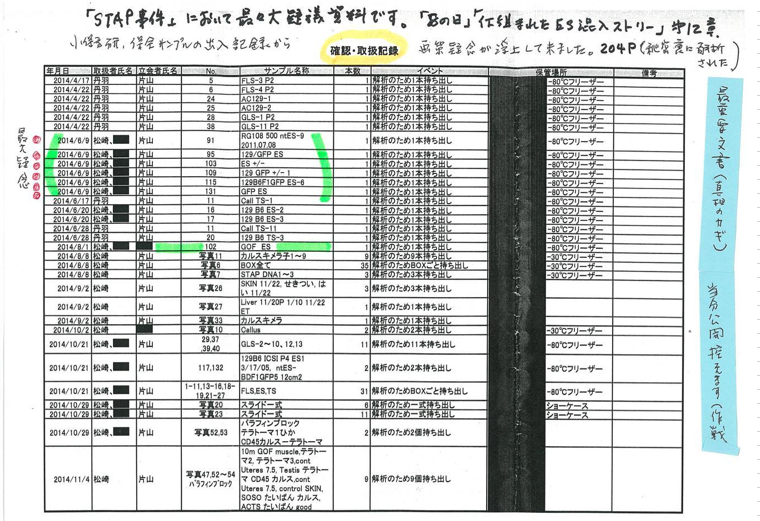
しかしこの時に全解析を受けた⑫はAC129の6番染色体にあった遺伝子異常と一致しなかった。そのため、ES1~5の6番染色体だけのシーケンスが行われたとみられる。このシーケンス結果が添付されてない理由はそれぞれがすべて微妙に違っていて、それを見せると一般の人たちから違うじゃないかと言われるからです。Extended Data Figure 2-cを再掲します。3度目の検討になります。
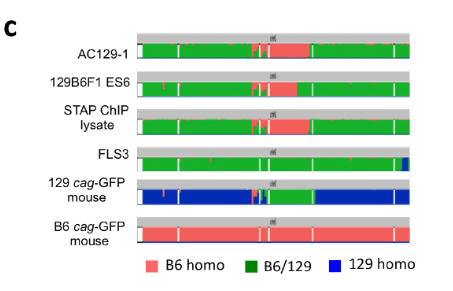
6番染色体です。B6マウスはすべてピンクです。129マウス中央にグリーン部分がある。ここはピンクとブルーが半々の場所です。ピンクとグリーンの掛け合わせからすべてグリーンの部分はできません。逆に言うとF1側でグリーンになっている場所は129側ではブルーだということです。
ここで分かることは上図に挙げられている129 cag-GFP mouseのSNPs分布はいろいろある129の中の一つに過ぎないということです。129のマウスコロニーは均一ではないのです。均一でないマウス同士の掛け合わせが行われて「僕のマウス」ESが作られている。均一だったら二種類のSNPs分布のESしかできない。それが1~6まですべてわずかに違っているんです。それは129マウス自体に何種類か混じっていてESが作られるとき複数のペアから採卵されている可能性もあるということです。桂報告書はこのことを以下のように説明している。9Pです。
>>
4)他の細胞株における遺伝的不均一性
この遺伝的不均一性は、129 CAG-GFP マウスに由来する他の細胞株にも反映していた。 若山氏により 129B6F1CAG-GFP マウスの独立した胚より複数の受精卵 ES 細胞株が樹立 されているが(129B6 F1ES1~6、2012 年 5 月作製)、いずれも第 6 染色体中程に B6 ホモ 領域を有していた。しかし、この B6 ホモ領域と 129/B6 ヘテロ領域の境界は 129B6 F1ES1 ~6 の間で異なっていた。このばらつきは、129 CAG-GFP マウスの配偶子が形成される 際、減数分裂の過程で、B6 と 129 の染色体の組換えによって生じた可能性が高いと考え られる。ただし、細胞株樹立時の体細胞分裂における染色体組換えがこの多様性に寄与 した可能性もある。ES1~6ですべて微妙に異なっていたことを認めています。素人に説明するのは大変だと見て意図的にシーケンス結果を並べなかったんです。
そして見せないままにその原因を減数分裂時と体細胞分裂時の染色体組み換えに求めている。しかし、そんなに都合よく微妙な場所だけで組み換えが起こるということは考えられない。
そもそも大きなヘテロ領域があると減数分裂で基本的に二種類できると先に説明したことを忘れている。SNPs分布でAC129 typeとES6 typeに分けましたよね。本当は2種類どころではなかったということです。
マウスコロニー自体に微妙な不均一が入っていて複数種のペアから採卵されているとこうなりますね。若山さんの維持している129CAGホモは長く維持しているからそれなりに近交系マウスになっていて維持メイティングはインクロスになっているはずだということを上述しています。途中で又B6が飛び込んでいるんですね。事故コンタミですね。気づいて慌てて取り出してもすでに交接してたということでしょう。無論、自家繁殖ですからこういうことはあるんで、実験の目的に支障なければ問題ないんです。若山さんの実験室では困るような事態では無かったということです。小保方さんの実験に使って、捏造事件になったから問題になっただけで、逆にマウスのコンタミが無かったら識別できなかったでしょうね。そもそも製薬の実験なんかに使われるマウスでこんなことがあったら大問題で、だから自家繁殖させてはいけない契約になってるわけです。若山さんはただ自分のところで使ってるだけです。まあ、予算節約ですね。まさか予算請求して懐に入れると飛んでも無いことになりますからそれはやってないでしょう。予実対比されますからね。経理部門がチェックしている。経理部門が何かしてるとまた別問題で、今度は会計監査が問題になる。そして公的部門には国税は入らないですからねえ。はは。
強引な結論に導こうとしてデータ開示しないからこういう訳の分からない説明になるわけです。cのリジェンドに以下のように書いていますが、書いている本人がクッソーと思いながら書いてるんじゃないですか。
>>
Note that the 129/B6-heterozygous SNP region in the 129 cag-GFP mouse is longer than that of AC129-1.
[ カツラ報告書 ]氏が指摘された場所もそれが原因です。
c.MTA無しの山梨大への移籍若山さんは2013年3月に理研内ラボを引き払うに際してMTAを締結しなかった。これは既に笹井さんが参加していてヴァカンティと特許の件に関しても話がついていて、米国側から守秘に関する依頼があったものと思われる。MTAを締結すると山梨大側にその情報が漏れてしまうということがあって、本来なら締結が必要であるが、論文発表まで待って事後締結すると打ち合わせられていたと推定される。手記に事務方から若山さんがMTAをなかなか出さないので訴えるという話を小保方さんが聞かされているのは、無論、イレギュラー処理をしていることを小保方さんに隠そうとしているので、若山さんとは打ち合わせ済のことである。こういう事務処理はマニュアル化されているので忘れるなどということはない。意図的に行われているのである。
しかし、このときにイレギュラー処理したがために、事件化後、理研は山梨大への若山ラボの冷凍サンプル保全命令依頼をできなかった。MTAも無いので山梨大はこのことを全く知らないからである。その結果、若山さんは理研の調査が始まっているというのに被疑者であるにも関わらず、自らが潔白であることが当然であるかのことくにサンプルを勝手に分析に出すという振る舞いができたのである。
2014/4/1に事後提出されたMTAの添付持ち出し資料にはコントロールESはES1しか持ってないことになっていて、3/10に放医研に出したコントロールESが5株であることは既述しています。調査体制の調整が何ら取られていないということも申し上げた。
この事後MTAに関して開示請求を行ったのが木星さんです。その後、このリストを使った持ち出し記録を開示請求したのはOoboeさんのパートナー氏です。
小保方さんが2012年8月にGRAS提出してLetter Figure 4-bで使ったESは以下の木星リストの131番ではないかと疑義されます。これはおそらく「僕のマウス」ESの1と6ではないか。

若山さん由来のGFP ESと言ったらコントロールの「僕のマウス」ESしかありません。小保方さんの略し方で129B6F1は129で、129ESと書いたらFLSのことでESはES-likeだと分かっている。
こういう書き方は無論FLSという名称が決まる前の状態でのサンプルです。STAP細胞が最初はスフィア→カルス→アニマルカルス→STAP細胞と変遷していくのと同じです。FLSという言葉が与えられる前はES様細胞と考えられているわけですからES-likeで、小保方さんはまだF1のコントロールESが作られていない時期ですから129ESと書いたら129B6F1のES-likeと自分で分かるからそう書いている。コントロールESを貰った後もGFP-ESと書いたらそれと分かる。学生から貰ったGOFのntESに関してはGOF-ES1と書かれていて、調査時に(■■由来→小保方)と答えていているのですぐに識別できているわけです。
でも、小保方さんは調査時には何が起きたのか全く分からない状態ですから、先生のラボに迷惑を掛けることもできない中で、口ごもりながらおどおどと答えることになる。調査側の考慮がまるで無い。周りが保身で汲々としている状態のようですね。こういう状態だと生贄の山羊になり易い。
GFP ESと書いてしかも若山さん由来だと、これは本当のES細胞です。これが1番と6番だったものを何度も解凍凍結を繰り返してラベルも書き換えられたものではないでしょうか。2本ある。旧DORAブログにある写真です。

これでは若山さんには気づけませんから若山さんは6は無いと思って、117番を忍び込ませたのではないか。松崎氏は131番を持ち出して調べているにも関わらず結果報告をしていない。
謎めいたところですが事実は以下です。
>>
理研小保方フリーザーにあるF1ESは以下です。
129 B6 ES-2
129 B6 ES-3
129 B6 ES-4
129 B6 ES-5
129B6F1GFP ES-6
GFP ES
若山さんが放医研に出したと言ってるF1ESは以下です。
129B6F1-1 ES
129B6F1-2 ES
129B6F1-3 ES
129B6F1-4 ES
129B6F1-6 ES
若山さんの事後MTA締結添付持ち出しリストにあるF1ESは以下です。
ES細胞 129B6F1 GFP-1
d.引っ越し時のサンプル整理上述した131番の下の132番はntESのBDF1ですから若山研で本来行われている実験のサンプルが紛れこんでいる。小保方さんは当然「不明」と答えています。
因みに若山さんの奥さんが警察の調べに自分のものと言ったと小保方さんが聞いたという日記の話はその132番と以下の117番です。それぞれ1本ずつの計2本です。

DORAさんの旧ブログに写真がある。GFP ESの2本も見えます。彼女の字です。若山さんのだとすぐわかったのだから自分の字なんです。このSの字は特徴がありました。P40 FLS-1~8のSです。彼女はちゃんとFLSの40継代120日間を実施しているんです。
小保方晴子日記は以下です。
>>
2016年
2月16日(火)
・・・私が新しく聞いた話もある。窃盗の疑いがあるというES細胞についての説明を受けた。報道された元留学生が作製したES細胞に加え、「ヒッポさんのもの2本」と写真を見せられた。その写真は理研の調査委員会の調査期間中、「持ち主不明のES細胞」と呼ばれていた細胞チューブだった。私は見知らぬES細胞が自分のサンプルボックスに入っていたことを知らされ、薄気味悪く思っていた。結局、「持ち主不明」と呼ばれたまま、調査は終了した。ともに公の場であるにもかかわらず、調査委員会で知り得なかった新事実を、事情聴取の場で聞かされた衝撃は大きかった。思わず刑事さんの話を遮って「ご本人の証言ですか?」と聞いてしまった。「そうです」と返事された。
ヒッポさんのだったんだ。盗まれたって言ったんだ。
ビニールとかプラステチックとか、そういう消化できないものを飲み込んでしまったみたいに、まさに消化できない気持ち悪さが残った。松崎氏は持ちだして調べています。報告がない。盗まれたと奥さん自身が警察の尋問で答えたのかどうかは日記の記載だけではわかりませせんが、石川氏がフライデーで盗まれたのだと書いたのは間違いありません。彼は山梨に訪ねて行って、若山さんと会ったが若山さんは体調が思わしくないので、奥さんと大日向氏が石川氏に説明したのでした。
因みに小保方さんの筆跡は以下です。日経サイエンスにこの写真が流出したのは調査に関与していた松崎氏のラボからです。P40のFLSが1~8までそろっているのに皆でスルーして、増殖率表の捏造だと騒いだんです。何なんでしょうか、この人たちって。
FLSのSの字を上のGFP ESのSの字と見比べると同じ筆跡と分かる。
40パッセージが終了したのは120日後の5月末頃ですから、その時にはすでにFLSという言葉が作られている。
 e.若山さんによる捏造
e.若山さんによる捏造このAC129とFLS-Tシリーズに関する捏造と他の捏造とされているもの(我々の説では岡部マウスとのF1マウス等を使った小保方酸浴細胞核使用の正規のntES実験)とは違うものです。
最初のキメラ成功から翌年初頭の確認実験や続く胎盤実験とCTSの作製までは、そもそも小保方酸浴細胞核使用ntESの性質解明実験で、実験自体は何も捏造実験ではなく、F1に関して使われているB6は最初からアクロシン入りの岡部マウスであっただけの話です。
太田ESや学生が渡したGOFのntESを小保方さんが捏造させるために若山さんに渡したのだという話は、若山さんが最初リクルート上の理由があって、小保方さんを引き留めるために、本当のことを言わずに、スタンダードな手法でキメラができたかのように小保方さんに話していて、ヴァカンティまで本気にさせてしまったので、いざというときには自分のESコンタミで言い訳しようとしてヘテロの話を造って、その伏線も小保方さんに吹き込んでいたものです。最初から小保方さんの所為にしようとして作った話ではなく、自分のミスだったと言い訳するための作り話に過ぎなかったものだ。無論実験自体は何も捏造ではなく、本当のことを小保方さんに言ってないがために論文が嘘になっていただけだ。若山さんは通らないと分かっていて、取り敢えず論文を書かせれば小保方さんはヴァカンティから自由になれると思っていた。それが本当のことを言わないままに小保方さんに論文を書かせていた理由です。小保方さんを理研が引き抜いた時から若山さんが事実を正直に言わない限り、いざというときのための言い訳として小保方さんの意図的コンタミというストーリーに変更せざるを得なくなった。道は3つに分かれていた。
①本当のことを言う(自分のプロモーションの話も含めて今更いえなくなってしまっていた。)
②論文のリジェクトを願う(専門家として通るはずはないと予測するのはそれほどおかしくない。そもそも三誌リジェクトされている。)
③小保方さんの所為にして逃げる(笹井さんの圧倒的な信用力で通ってしまいそうになり、現に最終的に通った。)
AC129の実験はまだ小保方さんが若山研に所属していて笹井さんが入る前の実験です。細胞の培養開始日は2012/8/13です。小保方さんは129/Svの赤ちゃんマウスを2012/8/5前後に若山さんから受け取ってSTAP細胞を作った。この実験がとても不可解なものでした。
f.混乱期の状況それを考える前に、この頃の状況を確認しておくと、小保方さんがプイと米国に帰ってしまったのは手記によれば、2012/11/15の数日前ですから2012/11/10前後ですが、小保方さんはこのAC129の実験が開始された2012/8/5頃の丁度3か月後にヴァカンティの許に帰ってしまった。つまり山梨にはいかないと返事したことになる。この返事の仕方は手記を読む限りでは、とても失礼なマナーです。ちゃんと筋を通すことなく、寺下さんにアメリカでやりたいと内心を打ち明けて渡米した。若山さんの耳に入ることを承知で打ち明けたでしょう。無論、渡米した以上、ヴァカンティや小島、大和に事前に相談している。そのことも手記に書いてある。しかし、推測では米国から小島を通して共同研究解除の話をしていると思われる。その後すぐに西川さんから理研への勧誘の話が入って、結局ちゃんと本人が挨拶してない形のままではなかったかと疑われる。直後に理研の話が入ったので気づかれにくいが、理研が誘わなかったら、小保方さんは若山さんとヴァカンティの間でヴァカンティを選んだという形になる。結構、この形は失礼でしょう。よほどちゃんと大和や小島と一緒に訪問して、自分の気持ちを説明し、お礼を言ってヴァカンティのところに戻るということをしないと、若山さんのプライドを痛く傷つけたと思います。日本とは学制が違うとはいえ、ヴァカンティは一介の麻酔医で博士号も取得していない人だ。方や、若山さんは世界初のマウスのクローン作製とntESの作製に成功している世界的な学者だ。記者会見で一度ヴァカンティと呼び捨てにして、後にヴァカンティ先生と言い直しています。特にリクルート絡みで対抗していましたからあまりしっくりした仲ではないでしょう。まあ、大人ですから態度にはださないでしょうが、結構ムッと来るような仕儀だと第三者が読んでいても感じるところですね。「残念だけれども山梨大の助教よりも理研のユニットリーダーを選ぶのは当然の選択です。面接頑張ってください」と励まされた若山さんの言葉を書き留めているんだけれども、理研が誘わなかったら、一介の麻酔医であるヴァカンティ研で一介のポスドクであることを選ぼうとしたのがあなたの当然の選択なのだろうかという皮肉を、小保方さんは感じたでしょう。若山研の客員に決まった経緯の中で小保方さんは若山さんに論文が出るまではヴァカンティ研に所属してくれと言われたと説明しています。手記にちゃんと書いてある。それはヴァカンティに言われたとおりのことなんでしょうけど、若山さんは論文が出たら彼女のリクルートは可能だとなんとなくは思っていたでしょう。そして、キメラは出来なかった。帰ろうとする彼女を翌年の山梨大助手の条件提示できるまでの時間稼ぎとして、別途考えていた酸浴細胞核を使ったntES化実験でできたキメラを説明なしでできたと一時的嘘をついた。そして翌年に助手で誘ったけれども彼女は即答しなかった。これでどんどんこじれて行ったんです。人はそれぞれ思いが違います。彼女は女性ですからあまり地位に拘りません。男性みたいな権力欲が薄い。米国の空気は自由なんで結局彼女がヴァカンティのもとに戻りたくなったのはそれが一番の原因だと思います。それに対して若山さんは米国の過酷な競争社会を知ってるし、父性から自分のところで落ち着いて成長させてやりたいという気持ちもあるんでしょう。こういうのってドモナランですなあ。世の中のことは誰でも自分の思い通りにはならない。それで運命という言葉もあるんでしょうや。
B. 欠失と重複a.全解析まずBCA報告のExtended Data Figure 2-c,dを再掲します。
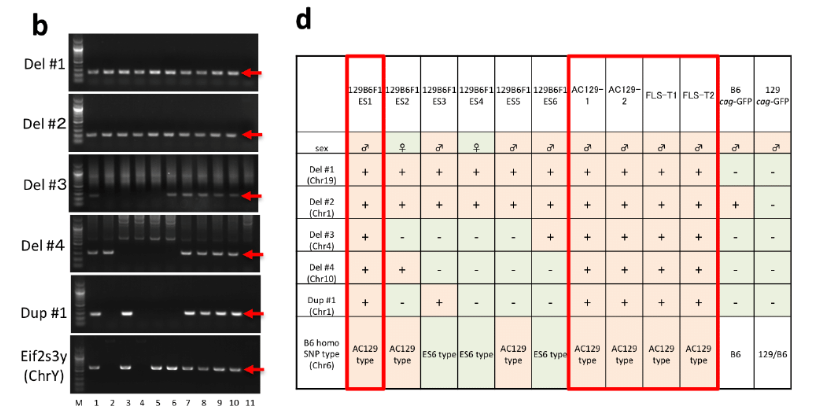
bとdのリジェンドは以下です。
>>
b, PCR detection of chromosomal anomalies and Y chromosome in cag-GFP STAP stem-cell lines and parental mouse strains. Lanes 1–6: control ES cells, 129B6F1 ES1–6; 7: STAP stem cell AC129-1; 8: STAP stem cell AC129-2; 9: STAP stem cell FLS-T1; 10: STAP stem cell FLS-T2; and 11: GOF-ES. Deletions 1–4 and duplication 1 are located on Chr19: 32,857093–32866,121, Chr1:140,698,249–140,702,693, Chr4:123,747,239–123,763,596, Chr10:43,265,147–43,267,270
d, Table summarizing the chromosomal anomalies and differential types of Chr6 B6-homozogous SNP clusters in the cag-GFP cell lines and parental mice. Control ES cell 129B6F1 ES1 shares all the characteristic features with the four cag-GFP STAP stem-cell lines.この中でWGSに掛けられた最初の細胞サンプルはAC129-1です。桂報告書の8P。
>>
小保方研フリーザーに保管されてい た STAP 幹細胞 AC129-1 について、SNPs マーカーの TaqMan PCR 法による解析を行い、さ らに NGS により全ゲノム DNA 配列を解析した。同じく STAP 幹細胞 FLS の対照として CAG-GFP マウスから作製された受精卵 ES 細胞(129B6 F1ES)の解析も同時に行った。まず最初にAC129が全解析されて、4個の遺伝子欠失と1個の重複が見つかった。BCA報告に貼付されているWGSに掛けられた細胞サンプルのリストは以下です。
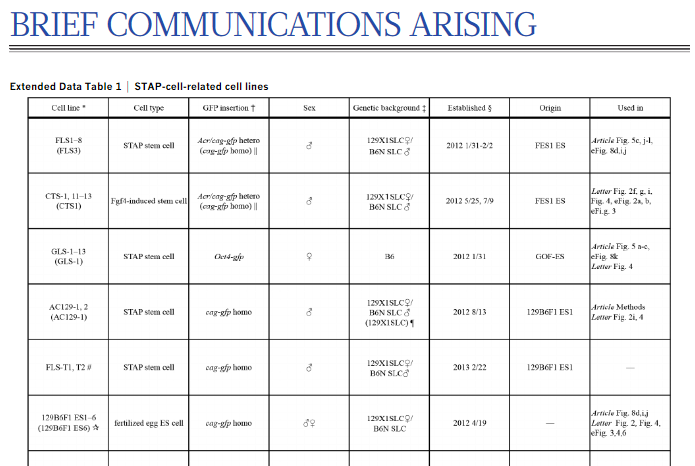
以下が関係したリジェンドです。
>>
*The line subjected to WGS is indicated in parentheses in cases in which several sublines were established for one cell type. Other sublines were confirmed by PCR and sequencing.>>
☆The genome of 129B6F1 ES6 was sequenced, but further analysis showed that 129B6F1 ES1 rather than 129B6F1 ES6 shares all genomic anomalies found in AC129-1.control ES 129B6F1 ES1はWGSには掛けられていません。掛けられているのは129B6F1 ES6で、他はOther sublines were confirmed by PCR and sequencing.とあるとおりです。でも、その結果は図示されていない。理由は既に述べた通り、全部違ってるからです。違うなら違う理由を説明すればいいだけですが、厳密にはわからなかったので、図は世間には見せないことにしてしまった。
それをごまかす為にb図の並びは4欠失と1重複、そして性別なのに、d図では性別を先にし、4欠失と1重複、そしてES1型とES6型の二種類のSNPsパターンに分類されるとした。図をつけると全部違うからつけなかったということです。ES1型をAC129型としているのは最初にAC129を全解析して、次にES6を全解析したが、遺伝子異常もSNPs分布も違っていたので、更にその後にES1~5の6番染色体だけをシーケンスしたためです。全解析されているのはAC129とES6だけです。AC129には4つの欠失と1つの重複があって他の染色体にはない。ES6には3つの欠失があって他の染色体にはない。ではES1~5の遺伝子異常はどうかというと全部は調べられてないということです。AC129の全解析で見つけられた以下の異常個所だけをPCRで確認したんです。
>>
①Del#1(Chr19)
②Del#2(Chr1)
③Del#3(Chr4)
④Del#4(Chr10)
⑤Dup#1(Chr1)
他の場所に有るか無いかはこの方法では分からない。ES1はPCRでAC129にあった遺伝子異常だけ調べて他は調べてないから同じだという証明は不完全ですが、結論は以下となっているのです。
>>
STAP 幹細胞 AC129-1、AC129-2、並びに、STAP 幹細胞 FLS-T1、FLS-T2 は、129B6 F1ES1 に由来すると結論づけた。 正しくは、現在のサンプルの中身はAC129-2、STAP 幹細胞 FLS-T1、FLS-T2 及び129B6 F1ES1 はSTAP 幹細胞 AC129-1の遺伝子異常と6番染色体上のB6と129のそれぞれの特徴的SNPs分布を
共有しているということです。
そもそも何がどう使われたかなんて、AC129-1の中身をAC129-2、STAP 幹細胞 FLS-T1、FLS-T2 及び129B6 F1ES1に入れてもそうなります。先にサンプルの証拠能力を調査検証してないから何事も言えません。全サンプルMTAもなく無断で若山さんは山梨に持ち出してるんですからどうにでもできる。
b.細胞の同定さて、まあ、それでもAC129-1の遺伝子異常は以下の4つのサンプルと共有されていたことは事実なんでしょう。
①129B6F1 ES1
②AC129-2
③FLS T1
④FLS T2
①のマウス背景は実物があって「僕のマウス」です。調べた結果分かっている。若山さんが一人で作って若山さんが持っていて、かつ小保方さんに渡したものもある。ただし、事件化後に残されていたサンプルの保管状況では小保方さんはES1をもって無くて、それは山梨で若山さんが持っていた。
②のマウス背景はAC129のCAGホモだと若山さんが証言しているし、白毛なので小保方さんも赤ちゃんマウスを渡されたときには1のアグーチとは違うと分かります。でも出来上がったものは「僕のマウス」だと調べがついてますから、若山さんが小保方さんの捏造と見せるために自分でマッチポンプの捏造をしたか、小保方さんが白毛のマウスを渡されながら、愚かにもひょっとしてキメラを作られたらすぐばれてしまうのに「僕のマウス」ESを渡したか、キメラは作らないと知らされていたので違う背景のESを渡したのどちらかです。
論文にはこの129/Svでキメラが作られたと書かれています。小保方さんが犯人だとこれは嘘になります。それと論文ではこの129/Svは129/Sv carrying Rosa26-gfp と書かれていて、これも小保方さんの嘘ということになってしまうんですけど、CAGホモと聞かされているか、もしくは何も知らされていないのにcarrying Rosa26-gfp と書くと、若山さんに嘘を指摘されるからそんなこと書かないでしょう。若山さんかもしくはラボの誰か、特に奥さんがそういったということで、この段階ではすでに小保方さんは嵌められようとしている。
もしくはそもそもこの実験では129/SvのCAGホモは使われていなくて実際には129/Sv carrying Rosa26-gfp だったのに若山さんが後から嘘にしているのかもしれない。この場合は理研も嘘に加担していることになる。というのも笹井さんや丹羽さんは129/Sv carrying Rosa26-gfp の記述に何の疑念も感じていません。理研にはそういうマウスはいろいろとあるし、丹羽さんも実験で使っている。ところが桂報告書は、「129/Sv carrying Rosa26-gfp マウスは理研 CDB に導入された記録や飼育記録はないことから、 これは誤記と考えられ、若山氏の説明によればここで言及された STAP 幹細胞は AC129 であった可能性が高い。」と書いている。
これは入手経路にマウス会社との裏ルートがある場合は本当のことは書けません。裏ルートというのは独法にありがちな余った予算を関連会社に商品を買ったことにして預ける。民間でもよく行われて国税に摘発される架空取引です。役所には税務署は入らない。これで予算を余らせないで済む。まあ、そういうところは蛇の道は蛇で、誰でも知ってるが、そんな話はここでの目的ではありません。
③④に関してはそもそもどんなマウスを小保方さんに渡したかはどこにも書かれていない。ただ、FLS-Tと書かれているから、FLSは「僕のマウス」を渡したと言ってるんだから、この場合もそうだろうと推測されているだけです。TはTeruで、若山さんの名前の照彦の略です。若山さんは一人で幹細胞まで樹立してそれを山梨大に持ち去ったわけですが、無論これは一人でできるかと試したものなので小保方さんには残していない。そして桂調査とBCA調査で129B6F1 ES1で捏造されていると証明されて、小保方さんが「僕のマウス」ESをインキュベーターの中でそっとコンタミしたのだという結論にさせようとしたのに、どれがどれと分からないと思っていたからでしょうが、ドジなことに若山さんは大本の129B6F1 ES1を山梨大に持ち去っていた。そして小保方さんのフリーザーボックスに129B6F1 ES6が残されていて、小保方さんは桂報告の公表前にこれに関して由来不明と答えているんです。警察だったらまず最初に疑われるのは若山さんです。
AC129の実験を行うとき若山さんはAC129のCAGホモを渡したと言ってるわけです。桂報告書には何らその証拠は提示されていません。FLSのときに「僕のマウス」を渡したという話も何の証拠も提示されていません。実験ノート記載くらいは見たんでしょうか。実験ノートにも書かれていなかったようですが。ただ若山さんがそういってるということです。呆れます。ノートに書かれているのなら実物を提示してくれないと困りますが、Ooboeさんのパートナー氏やDORAさんが開示請求しても応じません。以下はDORAさん資料です。

小保方さんが米国側の要請で開示できないということはさんざん非難したくせに。そして小保方さんが入院中に手元に置いていた実験ノートは強引に持ち帰り、全部コピーを取って、しかもNHKにその全コピーを違法に流出させた。何をやってるのか。何しろ今までごまかしばかりやってるから、彼らのやることは何事も鵜呑みにはできないと世間は見てます。
ただ、129/Svは白毛ですから小保方さんは白いのを受け取ったとは確認してます。若山さんはこの時キメラは作らなかったと証言してますが、小保方さんは作られたと論文に書いている。よくもまあ、こんなに食い違うものだね。若山さんは論文読んでないのでしょうか。
c.AC129実験の目的問題はAC129の実験とは何なのかということです。再掲しましょう。
>>
(d) STAP 幹細胞 AC129 は、129B6F1 マウスから作製された受精卵 ES 細胞に由来する
(調査結果) STAP 幹細胞株樹立に遺伝的背景が及ぼす影響を調べる実験が、若山氏により 2012 年 夏から秋にかけて行われ、2012 年 9 月 4 日に STAP 幹細胞 AC129-1 および AC129-2 が樹 立された。若山氏が交配した 129/Sv-CAG-GFP マウス(CAG-GFP 遺伝子が第 18 染色体に挿 入されたホモ接合体)由来の脾臓 CD45 陽性細胞を材料とし、小保方氏が STAP 細胞を作 製し、それを用いて若山氏が STAP 幹細胞として AC129 を樹立した。この細胞ストック はCDB若山研が山梨大へ移転する際に山梨大へ運ばれ、また一部は小保方研へ分与され、 それぞれフリーザーに保管されていた。このうち、小保方研フリーザーに保管されてい た STAP 幹細胞 AC129-1 について、SNPs マーカーの TaqMan PCR 法による解析を行い、さ らに NGS により全ゲノム DNA 配列を解析した。同じく STAP 幹細胞 FLS の対照として CAG-GFP マウスから作製された受精卵 ES 細胞(129B6 F1ES)の解析も同時に行った。得 られたデータを他の細胞等の解析結果や公開データと照合した結果、以下のことが判明 した。 「同じく STAP 幹細胞 FLS の対照として CAG-GFP マウスから作製された受精卵 ES 細胞(129B6 F1ES)の解析も同時に行った。」という書き方がレトリカルであるということがわかります。AC129-1が全解析されて、続けて「同じく」と書くとそちらも全解析されたかのように誤読されやすいのを分かってこう書いているんです。実際にはどういう解析なのか示されていないが、先に考察した通りです。示されているのは129B6 F1ES6の全解析結果だけなのに、それを含めて「(129B6 F1ES)の解析も同時に行った。」とラインをわざと書かずに紛らわしくさせているのです。報告書のスライド版も同じです。全解析したもののみのリストであるはずなのに129B6F1とだけ書かれていて、6つある中のES-6であってES-1ではないということが示されていない。意図的な隠蔽です。
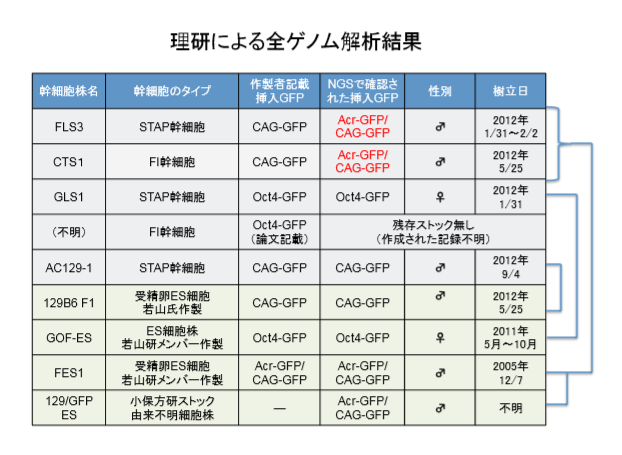
ともあれAC129の実験の目的は「STAP 幹細胞株樹立に遺伝的背景が及ぼす影響」だということです。
2012年の8月にもなってまだ「STAP 幹細胞株樹立に遺伝的背景が及ぼす影響を調べる実験」をしていたというんですが、2011年に既にF1とGOFで幹細胞化させています。最初のF1は「僕のマウス」ではないことは桂報告書が明らかにしている。翌年の初頭に又「僕のマウス」とGOFでやってる。B6GFP x DBA/2もやってます。その後はTCRの実験で又STAP細胞を作ってます。この時にキメラや幹細胞は以前のを使って、新たには作らなかったのでしょうか。更に胎盤が光ったという実験でSTAP細胞を作らせてキメラを作ってます。そのマウスの背景は何なんでしょうか。胎盤が光ってたんで最低でもCAGです。この時もSTAP細胞はリンパ球を使ってるはずです。コントロールの受精卵ESでもキメラを作って胎盤を小保方さんに渡しました。念のためにもう一回は再掲して置きましょう。
 若山さんはこの胎盤実験に関して左右の実験を一人で同時期に行ってることになるんです。これをやって酸浴リンパ球のナイフカットによるキメラ胚移植と、コントロールES細胞のキメラ胚移植で、細胞の大きさと、自分で引き延ばすパイプの太さが違うということに気づかない人はいません。
若山さんはこの胎盤実験に関して左右の実験を一人で同時期に行ってることになるんです。これをやって酸浴リンパ球のナイフカットによるキメラ胚移植と、コントロールES細胞のキメラ胚移植で、細胞の大きさと、自分で引き延ばすパイプの太さが違うということに気づかない人はいません。>>
(全録)STAP細胞論文の共著者・若山照彦教授会見 質疑応答(2/4)
8:30/42:12~
(朝日新聞岡崎女性記者)
で、先生は実際に、その、STAP細胞からキメラマウスを造ったり、その、幹細胞を作成されたりして、実際に、その、細胞を見てるわけですけど、見た目、外観からすると、全く今までと違った細胞で、あの、他の類似した細胞というのは見たことなかったのでしょうか。
(若山さん)
マイクロマニピュレーターの上に細胞を乗せてしまって、そこからキメラとかを作るわけですが、その状態になってしまうと、あと、あの、今回に関しては、それまでずっと失敗続きだったということもあって、ええ、普通、キメラマウスを作るときは、細胞をバラバラにして、キメラを作るんですが、塊のまま入れてみようという、そういうアイデアで実験をしたということもあり、いつもと違う手法を取り入れた、その時に成功したんです。そのために、その時の細胞が以前と違っていたかどうかというのが分からないままなんです。
(朝日新聞岡崎女性記者)
では、その、もしかしたら、その、万が一ES細胞だったとした場合、それを、これが新しい細胞ですと言われて、ほんとに、見分けがつかなかったんですかね。
(若山さん)
そうですね。その時点では、あの、新しい手法でやってしまったので、見慣れた外観は全くないので、どの細胞だったかということは区別できなかったと思います。嘘をついています。分からないことはありません。笹井さんも分かるはずだと言いました。若山さんはナイフカットは最初に一回やって写真撮っただけなんです。嘘ついてるんです。まあ、もはや我々はそれは前提でAC129の問題も考えています。
- 2019/08/27(火) 10:12:38|
- AC129
-
-
| コメント:0
『AC129を巡る問題』(1) AC129とは何であったのかA.経緯AC129-1,2とFLS-1,2は129B6F1ES-1によって捏造されたと桂報告及びBCA報告は主張している。
BCA報告のその部分は以下です。(参考 *ttps://www.researchgate.net/publication/283439572_STAP_cells_are_derived_from_ES_cells)
>>
Control genomic DNA sequences for STAP cell chromatin immunoprecipitation sequencing (ChIP-seq) experiments (Fig. 4 in ref. 2) had been deposited in the NCBI database(2). To gain sufficient sequencing coverage, we re-sequenced the genomic DNA prepared from the STAP cell lysate used for ChIP-seq (Extended Data Fig. 1a). We confirmed that this STAP cell sample shared all the genomic characteristics described above for 129B6F1 ES1 (Extended Data Fig. 2c),indicating that the STAP cell sample used for ChIP-seq was derived from 129B6F1 ES1 cells.ここで(Fig. 4 in ref. 2)と指定されているのは、Letter Figure 4-bのことです。
その図のSTAPとESのデータが入れ替わっているというのが若山さんによるリトラクション理由書にあるのはよく知られている。序ですから引用しておきましょう。
>>
(4) In Fig. 4b of the Letter, STAP cell and ES cell are wrongly labelled in a reverse manner.Letter Figure 4-bは以下です。よくこの程度の粗雑な簡略図でSTAPとESの取り違えに気づけたものです。誰かラボ内に手伝った人でもいて、元データを持っていた人がいたことが推察されます。
BCA報告の(Extended Data Fig. 2c)の図は以下です。
上表のSTAP ChIP lysateに関してBCA報告本文に説明がありません。書かれているのはNCBIのデータベースと、自分たちの分析図です。
>>
Control genomic DNA sequences for STAP cell chromatin immunoprecipitation sequencing (ChIP-seq) experiments (Fig. 4 in ref. 2) had been deposited in the NCBI database(2). To gain sufficient sequencing coverage, we re-sequenced the genomic DNA prepared from the STAP cell lysate used for ChIP-seq (Extended Data Fig. 1a). NCBIのデータベースに関しては、すでに我々が以前分類しているもので以下の分です。
>>
ChIP-Seq
⑩SRR1171582 小保方 Low pH treated CD45 positive Cells:ChIPSeq.H3K27me3 Oct3/4 expressing cells C57BL/6x129/Sv 129xB6-CAGヘテロ
⑪SRR1171583 小保方 Low pH treated CD45 positive Cells:ChIPSeq.H3K4me3 Oct3/4 expressing cells C57BL/6x129/Sv 129xB6-CAGヘテロ
⑫SRR1171584 小保方 Low pH treated CD45 positive Cells:ChIPSeq.input Oct3/4 expressing cells C57BL/6x129/Sv 129xB6-CAGヘテロこれがLetter Figure 4-bのSTAPの欄で上の緑がH3K4me3で下の赤がH3K27me3になっています。若山さんの取り消し理由書に書かれているのはESの分とSTAPの分が逆に提示されているという主張です。このデータはTs.Marker氏が再分析していますが、素人では見方が分からなくて入れ替わっているのかどうか確認できません。そのことに関する彼の見解もありません。
他方、⑫に関してデータだけでなく、混合液であるSTAP ChIP lysateが残っていたのは別の話ですから由来を説明しないと分かりませんが、BCA報告の(Extended Data Fig. 2c)の図は全く同じものが桂報告書のスライド版にも使われていて、本文16Pに以下のようにあるが、どういう形で保管されていたのかは説明されていない。
>>
3)STAP 細胞由来 ChIP-seq (input)サンプルは 129B6 F1ES1 から取得された
(調査結果) 小保方氏が CDB ゲノム資源解析ユニット(以下「GRAS」という)に持参し残されてい た STAP 細胞由来 ChIP-seq (input)サンプルを再度 NGS 解析した結果、STAP 細胞由来と される ChIP-seq input データは CAG-GFP の挿入を持つ 129xB6 へテロ系統由来の細胞か ら取得されたものと判明した。さらに SNPs の解析、特異的な欠失変異の解析により(2 -3-1-1(d)参照のこと)CAG-GFP が挿入された 129B6 F1ES1 とほぼ同一細胞由来の データであることが明らかとなった。2年前の生データがSTAP細胞だけ冷凍されて残っているというのも変で、残っているならこの時の試料の全てのはずですが、記者会見では伊藤氏が口頭で記者の質問に答えて、この時の分析の試料がGRASにたまたま残っていたのだと証言しただけで、他の試料の有無に関しての更なる追及も無かった。又その保管に関して文書で追加説明があったということもない。伊藤さんはBCA報告の著者の一人です。
>>
Daijiro Konno, Takeya Kasukawa, Kosuke Hashimoto, Takehiko Itoh, Taeko Suetsugu, Ikuo Miura, Shigeharu Wakana, Piero Carninci & Fumio Matsuzaki桂会見での古田女史の質問に対する答えは以下でした。言い終わった後の彼のしぐさと表情は精神分析学の研究対象に好例かもしれない。(*ttps://www.youtube.com/watch?v=M9oJGioHvIQ 1:15:00から)
>>
ただ、唯一やったのが、あ、GRASの方に残っていたChIP seq のinput 、しかもSTAP細胞のインプットはDNAとして残っていたので、それが10ページバワーポイントの、えーと、ところに書いてある、3番のアイテマイズして書いてあるところですけれども、これに関しては、えーと、30倍になるだけ、他のゲノムと同量となるまで読んでいます。そして、読んだ結果、この、同一であるということを、ま、認定したということで、それ以外に、それ以外に関しては、ま、残っていませんのでやってない、やってません。なぜ、たくさんある中の、唯一、STAP ChIP lysateだけが残されていたのか、どういう形で唯一残されていたのか、誰かの恣意で残されていたのか、それともGRASには残し方に関する規則は無いのか。裁判だったらまず最初に検討されるもので、ここにちゃんとした答えが無いと証拠採用はされない。
B.報告書の非論理と重大な疑念a.非論理まず、先にBCA報告のこの件に関するあからさまな嘘を指摘しておきましょう。
>>
We confirmed that this STAP cell sample shared all the genomic characteristics described above for 129B6F1 ES1 (Extended Data Fig. 2c),indicating that the STAP cell sample used for ChIP-seq was derived from 129B6F1 ES1 cells.上に貼り付けた(Extended Data Fig. 2c)には
129B6F1 ES1は存在していません。あるのはES6です。ES6を図示していながら、これもどこから持ってきたかも正式に記述しないSTAP ChIP lysateが、図に無いES1と同じSNPsパターンであることを確認したと書いているわけです。
東の空を指さしながら美しい夕焼けですねと言ってるような論理です。言ってる事実が少なくとも指さしている方向には無いので、言ってる事自体も本当か嘘か分からない。科学者が書いたとはとても思えない
非論理な文章です。
b.最大の疑念以下に示すのは木星リストですが、まず129B6F1 ES1はそもそも小保方フリーザーには存在していないものです。右端の書き込みは調査のために管理部署から持ち出された記録です。129 B6 ES-4と5には持ち出された記録がないということにも注意が必要でしょうね。PCRに掛けられたデータがありますから持ち出したことは確かのはずですが、記録をつけないまま杜撰に持ち出して調べている実態がうかがわれる


では129B6F1 ES1はどこにあったのか。以下の若山さんの事後MTA契約書添付の持ち出しリストの中にある。下から二段目です。
よりによって若山さんは129B6F1GFP-1を持ち出していました。AC129-1,2もFLS-T 1,2も若山さんが持ち出していたControl ES cell の 129B6F1GFP-1 で捏造されているとBCA報告は証明しているわけです。ところが桂報告書もBCA報告書もAC129とFLS-Tは129B6F1ES1によって捏造されたと声高に主張しながら、その試料がどこにあったかを一切書いてない。小保方さんのフリーザーにあったものではないどころか
若山さんが山梨大に持ち出していたのです。桂報告書もBCA報告も、AC129の実験は小保方さんが若山さんに129B6F1GFP-1を渡して捏造させたものと示唆しているのですが、使われた細胞は理研には無くて、山梨大にあったのです。そして、報告書はこんな重要な事実を報告していないのです。これが分かったのは木星さんを中心にした大勢の人々の公開資料の取り寄せ努力によるもので、理研自らが発表したことではありません。むしろ
彼らはそれを隠していたことになるのです。これが疑念の最初で最大のものです。
c.その他の疑念次に、桂報告とBCA報告の遺伝子解析によるAC129と129B6F1ES-1との一致証明に関する疑義です。これは以下の表で、桂報告書とBCA報告のどちらにも掲載されています。BCAのは遺伝子異常のPCR結果もついていますからそれを貼り付けます。
この表にある129B6F1 ES4と5には持ち出された記録がないということと、ES1が山梨の若山さんのフリーザーにあったということが書かれていないということは既に指摘した通りです。通常の事件ですと、こういう情報が揃うとまず最初に疑われなければならないのは小保方さんではなくて若山さんの方なんですけどね。先にこの所謂「僕のマウス」ESのサンプル保管の状況から検討しましょう。
(サンプルの状況)これに関しては既に見てきた通り二通りのラベル表記がある。若山さんの持ち出しリストに嘘か誠か6ライン樹立したと書かれています。嘘か誠かというのは、TSに関しては2ラインになってるが実際には129 B6 TS-3,4,5,6が理研側に残されていますからこれも6ラインのようです。嘘か勘違いかは問わず、今はESに関してのみ考えましょう。想定されるもともとあったラインは以下です。
①129 B6 ES-1,2,3,4,5,6
②129B6F1 GFP ES-1,2,3,4,5,6
ここで重要なのは今桂報告として知られている調査結果が世間に知られる以前の聞き取り調査段階で、小保方さんが①は若山さんが作ったと証言しているのに、離れた場所に入っていた②の6に関して「由来不明」と証言していることです。
つまり、小保方さんが知っているラベルの書き方は①であったということです。そして、②の1は若山さんが持っていて、②の6が誰かの置忘れか、意図的にそこに置かれたものだということになる。現状は以下です。
>>
ES細胞 129B6F1 GFP-1(若山持ち出しリスト)
129 B6 ES-2(小保方さん「若山TL樹立」証言)
129 B6 ES-3(小保方さん「若山TL樹立」証言)
129 B6 ES-4(小保方さん「若山TL樹立」証言)
129 B6 ES-5(小保方さん「若山TL樹立」証言)
129B6F1 GFP ES-6(小保方さん「由来不明」証言)
2013年の3月の引っ越し時に若山さんによるサンプルの整理が行われている。そして山梨大から放医研にサンプル調査に出したのは1年後の2014年の3月です。そして4月になってMTAの持ち出しリストに自分は現在1しか持ってないように書いた。
彼は記者会見で放医研に出した129B6F1 ESを5株と言っています。なぜ6株では無いのか。
そして、この5株は放医研側のPCR結果のデータには1,2,3,4,6と書かれている。5がなぜ提出されなかったのかの理由説明もない。
更にMTAの持ち出しリストには6株樹立されて、今自分が持っているのは1だけのように記載されている。放医研には株分けして出したのであれば少なくとも1,2,3,4,6が持ち出しリスト上に無ければならない。仮に株分けせず手持ちのものをそのまま出したのなら、放医研に出したものは記載しないのなら何もないことになる。1だけがあるという理由は1だけは株分け分があったということになるしかない。
ところがこのESは理研側でも調査されていているからにはそのときには全ラインあったということになる。理研側の調査したサンプルの由来はどうなっているのか。
小保方さんのフリーザーに残されていたものとその松崎氏の持ち出し状況は以下です。
>>
129 B6 ES-2 (2014/6/20 松崎氏持ち出し記録あり)
129 B6 ES-3 (2014/6/20 松崎氏持ち出し記録あり)
129 B6 ES-4
129 B6 ES-5
129B6F1GFP ES-6(2014/6/9 松崎氏持ち出し記録あり)
若山さんの記者会見スライドにあった放医研の記載の仕方は以下でした。ラベルは全部新たに書き換えられて提出されていることが分かります。
>>
129B6F1-1 ES
129B6F1-2 ES
129B6F1-3 ES
129B6F1-4 ES
129B6F1-6 ES
放医研のサンプルを回して貰ったのなら5がありませんが、どこから持ってきたのでしょうかね。129 B6 ES-5 は少なくとも持ち出し管理記録上は持ち出されていないことになっている。調査が如何に杜撰で恣意的かということです。
ただし、1~6は欠失重複等が全部違っていますから理研側が6種類揃えたことは間違いありません。何かの中身が重複しているということはありません。調査細胞の入手経緯が明らかになっていないだけです。
我々の推測では[129B6F1 GFP ES-6(小保方さん「由来不明」証言)]を小保方さんのボックスに置いたのは若山さんだと推測しています。若山さんは1を置いておいても6を置いておいても大して違わないと思っていた。というのも遺伝子異常があって全ライン識別されるとは思ってもいなかったでしょうし、雌雄の違いも考えている暇がなかったんでしょうね。1も6もたまたま雄でしたが、違ってたら言い訳のつかないことになってたでしょう。核型解析に出せばすぐ分かりますから事前に雌雄確認をしていたかもしれません。ただ、遺伝子異常まではさしもの若山さんも気づかなかったでしょう。ES-1は無論、直前にFLS-T1,T2を作っていた時に使った株分け分が手元に残ったのではないか。これが致命傷になった。これ見よがしに置かれていたES-6は使われていなくて、若山さんが山梨に持ち出していたES-1が使われていたという藪蛇になってしまった。
問題は理研側に残されていた分で129 B6 ES-1と129 B6 ES-6がなぜ無いのかということです。このコントロールESは小保方さんは論文に使っていますから、実際に使用している。別名でどこかにあるかもしれません。後に詳しく調べてみましょう。
(SNPs解析)SNPsでAC129タイプと書かれているのはES1タイプということです。ES1タイプのマウスとES6タイプの2タイプの卵が使われているということです。なぜこういうことになるのかを考えてみる。
SNPは一塩基性多型と呼ばれている突然変異で人間でもそれぞれの人の顔が違うというような表現型の多様性等を作っているものですが、無論人間であることは共通ですから大半の遺伝子は共通です。
一方近交系マウスは20世代以上の兄妹交配を重ねて相同染色体の同一遺伝子座の遺伝子をホモにそろえてありますからSNPsもホモです。近交系マウスはマウス業者(ラボ)が作って系統立てて管理しているわけですが、例えば129/SvJ独特のSNPs分布というのは最初にジャクソン研究所で作ったマウスのSNPsを引き継いでいるものです。B6のラインを作るときは又別のマウスを使いますからこれは亜種が違うので当然全然違う分布になる。しかし、129でもB6でも最初に使われたマウスの個性が引き継がれるので仮に別の人が一から作ると別のSNPs分布になる。それでは研究目的に使えないので、同じ名前にはなりません。ですから例えば129/SvJと言ったら最初にジャクソン研究所の作ったマウスと決まっていてそれが国際的にも引き継がれていて、SNPs分布も決まっている。
継代繁殖で新たにできるSNPsというのはわずかなので、長く蓄積すると更に別の表記を加えますが、直接的には関係ないんです。
で、SNPsパターンでES1タイプとES6タイプの二つあるというのは、雑種交配させたんですから、129とB6の混ざり方の違う2個の受精卵が使われているということになる。完全な近交系マウスでは減数分裂時にできてくる卵も精子もそれぞれに遺伝子配列は全く同じですからこんなことになることはない。従って一方が完全な近交系マウスになってないということです。その原因は所謂「僕のマウス」作成の経緯にある。
若山さんはロックフェラー大学にいるときにB6のCAGホモマウスを作った。日本に戻ってからそれを129と掛け合わせてその後戻し交配を続けてGFPだけを129に移し替えた。129もB6も無論近交系で、マウス生産業者が20世代以上の兄妹交配を続けて対立遺伝子をホモにそろえて販売しているものです。契約があって原則自家繁殖させてはいけない。特に人にあげてはいけないです。マウス業者がその純度を保証できないから予期せぬところでの責任を負いきれないというのが理由です。
最初にB6の卵にウイルスベクターを使ってCAG-GFPを挿入する。どこに入ったかは分からないが、入ったら生まれたマウスは蛍光する。この蛍光している一匹のマウスに又B6を掛け合わせてたくさん子供を作る。この時の子マウスのCAG-GFPが光っている雄雌だけをメイティングする。GFPヘテロ同士の掛け合わせになります。ヘテロ同士を掛け合わせると第3世代で1/4の確率でホモができる。ランダムに交配させて十分な子マウスを作って一匹もGFP無しの子マウスが生まれてこなかったペアはホモ同士という確認で継代を重ねることを省略できる。もともとが近交系マウスですからGFP以外は全部ホモです。そういうことをしないで念を入れて最初から20世代継代してもホモになりますね。(放医研の検査で18番にホモに入っていました。)
それに対して129に移し替えるというのはよりテクニカルです。ただ単に129のCAGマウスを作るだけならB6と同じやり方でできる。でもこれだと又GFPがどこに入るか分からない。GFPが染色体の同じ場所にホモで入っているB6と129が作りたいということです。
それでまず一匹のB6-CAGホモマウスと一匹の普通の129とを交配させた。これは最初129B6F1です。B6がGFPホモですから子供には全部GFPがヘテロに来る。ただし、雑種にしましたから他の遺伝子もヘテロに変わってしまっています。
そこで129への戻し交配を行って、GFP遺伝子以外を親の129の近交系に戻す。そのやり方はGFP蛍光のある子供に対してのみ戻し交配を行うんです。これを繰り返すとGFPのみヘテロの129近交系マウスに戻ります。このヘテロ同士を掛け合わせて129GFPホモを作るんですが、この戻し交配が完全に行われていないとGFP以外の他の遺伝子にもB6の遺伝子が残ってしまう。それが上に貼り付けているBCA報告のExtended Data Figure 2-cのSNPs分析です。再掲しておきましょう。
上図のリジェンドです。
>>
c, Distribution of B6-type and 129-type SNPs along chromosome 6. The B6-homozygous SNP cluster(magenta) in the middle, which probably arose from the inheritance of the parental 129, is heterogeneous in length among six control ES cell lines. The four cag-GFP STAP stem-cell lines share the same length of the B6 SNP cluster with control ES 129B6F1 ES1. Note that the 129/B6-heterozygous SNP region in the 129 cag-GFP mouse is longer than that of AC129-1.下二つが今説明した親マウスの6番染色体のSNPs分布図です。B6は全部ピンク(magenta) です。これはロックフェラー大学で作ったGFPマウスです。GFPを挿入しただけですから対立遺伝子は全く同じで全部ホモにそろっていますから問題ない。対して129の方は全部ブルーになってないといけませんが、ピンクの部分とグリーンの部分が残ってます。ピンクの部分は相同染色体の両方がB6で、グリーンは一方がB6で他方が129ということです。戻し交配の回数が不足している状態でGFPヘテロ同士を掛け合わせてホモにしてしまった結果かもしれません。因みにGFPホモであるかどうかは更にもう一代子供を作って、全部蛍光しているかどうかで確認できます。
問題は129です。近交系マウスに戻っていない。厳密にいうとGFPだけでなく、129/Svだということすらできない状態です。もう少し長くバッククロスを続けてから兄妹交配させないといけなかったのかもしれない。でも、調べない限り本人は気づいていませんからやり直すことはできない。できたものの兄妹交配を続けるだけです。しかし、これはこれなりに20世代交配させていたら近交系マウスになります。
マウスは20日で生まれて更に妊娠可能になるまで50日かかります。70日の20継代は1400日です。3.8年です。若山さんは日本に帰ってきてから129へのGFP移し替えを行いましたから、戻し交配でも3.8年かかっている。しかし、その後の20継代維持を入れると計7.6年ですが、理研の10年目だったわけですから、ちゃんと継代できていたら、全部ホモにそろっていたはずです。にもかかわらずヘテロ領域がこんなにあるというのはもっと近い時期にB6とのコンタミがあったということになるのではないでしょうか。又そもそもの129には歴史的にジャクソン研究所で一度B6が混入したという事件がありました。従ってピンク部分はもともと存在していたものかもしれません。
そもそも論としてこういう図形がどうしてできるかということですが、ひとつづつの対立遺伝子のあり方はホモかヘテロしかありません。大多数のDNAはマウスである限り共通ですから、129かB6のどちらということはできません。言えるのはSNPのある個所だけです。
このケースの場合で、6番染色体のDNAの並びをある一定区間で区切って1単位とします。この長さは適当に人が決める。この区間にSNPsが何もなければ白抜きで表示する。遺伝子のあるところは大事なところなんでSNPsは少ない。そういう場所は白抜きになる。存在しているSNPsがB6特異的な変異だったらピンクで表示する。129特異的な変異だったらブルーで表示する。半々だったらグリーンです。F1ですから父と母とから来た両方があるのが普通です。だから大半グリーンになる。
では混じっているが半々でないときはどうするか。半々の部分とB6が勝ってる部分、129が勝っている部分と1単位の中で棒グラフとして表示している。
1単位の区間を細かくすると例えば以下のBCA報告のExtended Data Figure 1-c図のようになるし、区間を広くとると上のExtended Data Fig. 2cのようなものになるわけです。
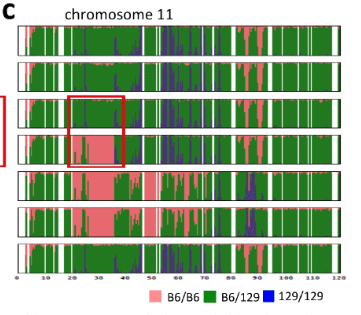
ともあれ、母親の129の染色体の一部にヘテロ部分があったわけですから卵原細胞が減数分裂するとき別々のSNPs分布を持つ二種類の卵ができる。マウスは多産ですのでたくさんの受精卵ができて、胚盤胞にまで成長させてから中のインナーセルマスを取り出してES細胞を培養する。8株できたわけです。それがES1型とES6型に分かれたのはそういう理由です。雌雄は以下です。
>>
ES1タイプ→ES1(オス)、ES2(メス)、ES5(オス)
ES6タイプ→ES3(オス)、ES4(メス)、ES6(オス)
この解析結果の不可思議はES6以外のSNPs解析データが図示されていないことです。このことは後に述べます。
(関連細胞)Letter Figure 4-bで使われている各種細胞は以下です。
>>
①CD45
②STAP
③ES
④STAP-SC
⑤FI-SC
⑥TS
Letter Figure 4-bそのものはこれです。再掲します。
この図に対応しているのが公共データベースに登録されている以下のデータです。
>>
ChIP-Seq
①SRR1171553 小保方 CD45 positive Cells:ChIPSeq.H3K27me3 derived from spleen Oct3/4::gfp C57BL/6 GOF
②SRR1171554 小保方 CD45 positive Cells:ChIPSeq.H3K4me3 derived from spleen Oct3/4::gfp C57BL/6 GOF
③SRR1171555 小保方 CD45 positive Cells:ChIPSeq.input derived from spleen Oct3/4::gfp C57BL/6 GOF
④SRR1171562 若山 Embryonic Stem Cells:ChIPSeq.H3K27me3 C57BL/6x129/Sv
⑤SRR1171563 若山 Embryonic Stem Cells:ChIPSeq.H3K4me3 C57BL/6x129/Sv
⑥SRR1171564 若山 Embryonic Stem Cells:ChIPSeq.input C57BL/6x129/Sv
⑦SRR1171567 若山 FI-SC(Fgf Induced Stem Cells):ChIPSeq.H3K27me3 Oct3/4 expressing cells C57BL/6x129/Sv 129xB6-Acr-CAG
⑧SRR1171568 若山 FI-SC(Fgf Induced Stem Cells):ChIPSeq.H3K4me3 Oct3/4 expressing cells C57BL/6x129/Sv 129xB6-Acr-CAG
⑨SRR1171569 若山 FI-SC(Fgf Induced Stem Cells):ChIPSeq.input Oct3/4 expressing cells C57BL/6x129/Sv 129xB6-Acr-CAG
⑩SRR1171582 小保方 Low pH treated CD45 positive Cells:ChIPSeq.H3K27me3 Oct3/4 expressing cells C57BL/6x129/Sv 129xB6-CAG
⑪SRR1171583 小保方 Low pH treated CD45 positive Cells:ChIPSeq.H3K4me3 Oct3/4 expressing cells C57BL/6x129/Sv 129xB6-CAG
⑫SRR1171584 小保方 Low pH treated CD45 positive Cells:ChIPSeq.input Oct3/4 expressing cells C57BL/6x129/Sv 129xB6-CAG
⑬SRR1171587 若山 STAP-SC(STAP derived Stem Cells):ChIPSeq.H3K27me3 Oct3/4 expressing cells C57BL/6x129/Sv 129xB6-CAG
⑭SRR1171588 若山 STAP-SC(STAP derived Stem Cells):ChIPSeq.H3K4me3 Oct3/4 expressing cells C57BL/6x129/Sv 129xB6-CAG
⑮SRR1171589 若山 STAP-SC(STAP derived Stem Cells):ChIPSeq.input Oct3/4 expressing cells C57BL/6x129/Sv 129xB6-CAG
⑯SRR1171592 丹羽研 Trophoblast Stem Cells:ChIPSeq.H3K27me3 CD1
⑰SRR1171593 丹羽研 Trophoblast Stem Cells:ChIPSeq.H3K4me3 CD1
⑱SRR1171594 丹羽研 Trophoblast Stem Cells:ChIPSeq.input CD1最初に注意しておかなければなりませんが、これらの試料がGRASに提出されたのは2012年の8月です。従って細胞の製作者は基本小保方さんと若山さんです。8月の解析は笹井さん参加前の実験結果で、しかも幹細胞化実験ですから、3誌論文の中には取り入れられてないものです。手記によると8月になってから若山さんが幹細胞化実験の成果を自分に51%割合で特許申請手続きを開始したとされている。そこから米国側と揉めて11月には小保方さんが米国に引き上げようとした経緯につながっている。
笹井さんが引き継いでからNGS解析をつけようということになってこの8月のデータが検討された。ChIP seqのデータもこのときにLetter Figure 4-bに纏められた。
若山さんは多忙で手伝えないということですから、新たに幹細胞やキメラを作ることはできない。公開データは笹井さん参加後にレター論文に使われたものです。若山さんは手伝っていませんから、若山さんが作ったものは2012年8月以前のものです。ES細胞とFI幹細胞は若山さんしか作れない。STAP幹細胞は小保方さんがSTAP細胞を作って若山さんがそれを使って作成した。CD45細胞とSTAP細胞は小保方さんはマウスさえあれば何時でも作れる。TS細胞は若山さんしか作れませんが、若山さんはF1で作ったと言ってますから、CD1マウス背景のTSは丹羽研のものと分かる。このことは若山さんの作ったTSが分化しているようだったので丹羽さんが提供したと桂報告書にも書かれています。桂報告書26P。
>>
TSについては細胞培養時に分化し たことが考えられたことから、丹羽研のメンバーが培養・サンプル調製を行ったものを 追加して作図に用いた。⑯⑰⑱は丹羽研のものと分かっているわけです。論文の論旨としては比較のためのコントロールですから誰のTSでもいい。2012年8月に提出されていたもともとの細胞はF1背景のTS細胞だった。
①②③は白血球ですが、GOFだと書かれている。これも論文ではGOFとF1が使われているわけですが、どちらでもいい。ただし、2012年8月のGRAS提出目的はレター論文用ではありません。笹井さんはまだ参加してない時期です。特許申請手続きを始めていますから、若山さん主導の幹細胞論文や特許申請書に添付するデータとして使う目的です。この時にGOFであったかどうかは分かりません。もしかしたら他と同じF1の分であったかもしれない。
その他は2012年8月のときのままです。酸浴細胞は小保方さんは何時でも作れますが、若山研のF1マウスは笹井研時代には使えませんから当時のままだと分かる。そしてすべての試料はF1背景です。ただし、理研が2012/2/13にNCBIに提出したときには今見られるとおりのC57BL/6x129/Svになっていた。しかし、小保方さんが理研に提出した2012/11/3の時点でどう書かれていたかは分かっていません。ここには若山さんの作ったES細胞があってそのマウス背景は129/SvxC57BL/6だということは分かっていますから、この登録の少なくともこの分は間違っている。ただ、それを小保方さんが間違えたのかは分かりません。
小保方さんはNCBIに直接登録はしていない。行ったのは理研の別人です。ここではF1であるという事にとどめておきます。
④⑤⑥がそのES細胞です。
⑦⑧⑨はFI幹細胞ですが、後の調査でアクロシンが出た。桂報告では太田ESだということになっていますが、若山さんは細胞の大きさで気づかないということはあり得ないと証明済ですので、これは岡部マウスとのF1由来の小保方細胞核使用ntESをTS培地誘導したものと分かる。
⑩⑪⑫がSTAP細胞ですが、これはアクロシンが出ていない。しかもCAG-GFPです。129でもB6でもCAG-GFPの入っているマウスは「僕のマウス」の親しか知られていない。小保方さんは見かけでは親の雌雄とGFPがホモかヘテロかはわかりません。
⑬⑭⑮が上の⑩⑪⑫をSTAP幹細胞誘導したものということになるが、これも後の調査でCAG-GFPと判明している。我々の判断ではこれもntES化しているということになるが、胎盤が光ると言われたキメラは小保方さんはB6/129としていますから、その時にSTAP幹細胞が作られていると知らされているのかもしれない。いずれにせよ。FLSは「僕のマウス」を渡したが中身は太田ESだったとされているが、
ここに公共データ登録では背景がB6x129と記載されている細胞で、事実中身が「僕のマウス」から作られているSTAP幹細胞と称する細胞があるということは驚きで、このことに誰も気づいていません。もしこれが「僕のマウス」ESの1~6のどれかなのなら、今でもデータはあるのですから調べたらわかります。しかし、SNPs解析も遺伝子異常解析もされていません。ただ、マウス背景が129B6F1のCAGホモだと桂報告が言ってるだけです。
Letter Figure 4-bは、ES細胞に代表される多能性遺伝子(Pluripotency marker genes)であるOct4,Nanog,Sox2、二価クロマチン(Bivatent pattern genes)であるGata2,brachyury,Nkx6-2、そしてTS細胞特異的遺伝子(Trophoblast maker genes)であるCdx2,Enomes Itgα7に関して、それぞれの、その巻き取られているヒストン領域がH3K4me3(H3ヒストンのK4サイトのリジンの3つあるアミノ基が全部メチル基に置き換わっているトリメチル化と呼ばれる現象を起こしている)とH3K27me3の状態にあるDNAをクロマチン免疫沈降(Chip=chromatin immunoprecipitation)で抽出した後に定量PCRに掛けた結果である。
(H3K27me3)は常時転写抑制として働き、(H3K4me3)は活性化を許可する働きを持つ。二価クロマチンは間に立ってその両方をコントロールする。従って例えばOct4遺伝子のCD45を見ると緑の(H3K4me3)状態にあるものはなく、赤の(H3K27me3)状態にあるDNA断片がたくさんあることを示している。その下のSTAP細胞を見ると赤はなく緑がたくさんある。つまりOct4遺伝子のプロモーター領域が活性化されているので結果的にはOct4遺伝子を発現させてOct4蛋白質をたくさん産出しているということになる。この状態でOct4-GFPが組み込まれていると細胞は蛍光することになる。
ChIP seqに限らず、この遺伝子解析結果を重視する人々、Ts.Markerさん、DORAさん、学さん、和モガさん、そしてOoboeさんとそのパートナーさんが"STAP有る派"となっていることは衆知の事です。そして彼らは同時にキメラはナイフ切り分けであれ、スタンダードなやり方で論文通りに出来ていると考えている。若山さん本人が太田ESのコンタミだと言って明らかな嘘をついて論文を否定した理由は何らかの陰謀であると考えておられるらしいが、それがどういうものかに関して何ら推論も論証も無い。
対して小保方細胞核使用ntES論は説明した通りである。
1.キメラはntESでできている。
2.幹細胞もntESでできている。
3.STAP酸浴細胞は小保方さんが作った本物である。
丹羽論文はティシュー論文の一部を再現したが、理研での酸浴Oct4-GFP蛍光細胞が本物のOct4遺伝子をほんのわずかしか含まず、GFP蛍な光の漏れ出し現象を指摘した。それに対して、このLetter Figure 4-bの実験結果はたくさんの内在性Oct4遺伝子の発現を定量的に立証している。つまり小保方さんの再現実験があまりに過酷で十分な再現条件でなかった可能性を示している。
以下は彼女の再現実験の予備実験時の写真です。旧DORAブログにあるものですが、近々閉鎖になりますから、あちこちに証拠保存しておくという意味でも貼り付けて置きましょう。参考までに、後にOoboe さんのパートナーさんの取り寄せた、赤色フィルター切り替えで自家蛍光の無いことの確認されている、全てが、緑色スフィア塊の写真も西岡さんのガンバレブログに楠本さんが貼り付けています。
しかし、だからと言って、キメラができたことにはならない。
さて、一方で、レター論文にNBCI登録されていると書かれているのは以下の分ですが、上述したChIP seq分、或はここにも書かれているChIP-seq files はここに挙げられているリストにはない。
>>
RNA-seq and ChIP-seq files have been submitted to the NCBI BioSample databases under accessions SAMN02393426, SAMN02393427, SAMN02393428, SAMN02393429, SAMN02393430, SAMN02393431, SAMN02393432, SAMN02393433, SAMN02393434 and SAMN02393435.このリストに対応している登録サンプルは以下です。
こちらのRNA分析にはES細胞が二つ登録されています。恐らく若山さんの作った分と、レター論文で使われたBFPのES細胞ではないかと思われます。これは丹羽さんが作っているのではないか。何にも確認されていません。ほんとに杜撰な調査です。すべて小保方さんや丹羽さんに聞いたら分かる事ばかりです。
報告書は二年前の2012年8月にGRASに提出されたLetter Figure 4-bのSTAPのグラフのもととなった上記⑫の生資料が保存されていたと説明しました。桂報告書16Pです。
>>
3)STAP 細胞由来 ChIP-seq (input)サンプルは 129B6 F1ES1 から取得された
(調査結果) 小保方氏が CDB ゲノム資源解析ユニット(以下「GRAS」という)に持参し残されてい た STAP 細胞由来 ChIP-seq (input)サンプルを再度 NGS 解析した結果、STAP 細胞由来と される ChIP-seq input データは CAG-GFP の挿入を持つ 129xB6 へテロ系統由来の細胞か ら取得されたものと判明した。さらに SNPs の解析、特異的な欠失変異の解析により(2 -3-1-1(d)参照のこと)CAG-GFP が挿入された 129B6 F1ES1 とほぼ同一細胞由来の データであることが明らかとなった。 ⑫があるにも関わらず、調査チームは保存されていたinputデータの混合液をもう一度解析したというのです。公共データ登録されているデータとどう違うかの説明はありません。inputデータは提出された細胞のDNA、RNAの全てを含んだ混合液のシーケンスデータです。その混合液自体が残されていて、これを他のゲノムと同量となる30倍になるまで読んだという伊藤さんの説明ですが、ど素人なのでその意味は分からない。
このSNPsの解析表がBCA報告の(Extended Data Fig. 2c)ですが、129B6F1 ES1が無いということは既に指摘しています。129B6F1 ES6とSTAP ChIP lysateはど素人が見ても全くパターンが違います。一方、AC129-1は129B6F1 ES1と同じだと言われているんですが、そのAC129-1とSTAP ChIP lysateとの比較はどうかというと、これはとても初期段階でTs.Marker さんが問題提起しました。
Ts.Marker さんは後にグラフィックスの精度の問題の範囲内かもしれないとおっしゃってますが、本日(2019/8/14)学ブログに以下の書き込みがありました。以前から指摘されていることですが、1Mbの範囲内で半分ほど違ってると改めて指摘されると、確かにこれは大きいですね。今のところSTAP ChIP lysate≠AC129-1(≒?129B6F1 ES1)ということではないでしょうか。
>>
拡張データ2CのSNPパターンの図ですが、縦の1枠(白抜きになっている部分)が1Mb(塩基配列100万個)です。縦枠150個並べて第6染色体の150Mbの全塩基を表しています。ChIP-seqとAC129は67Mb~68Mbの1Mbの範囲に違いがあり、AC129は全てB6ホモだけど、ChIP-seqは約500kbがB6ホモ、残りが129/B6ヘテロになっています。FLSの欠失は8番染色体で17kb、3番染色体の欠失は5kbで同じ欠失があることでFES1他と同一認定されましたが、違いが500Kbもあるというのは大きな違いです。細胞株が違うことを証明するのは、違いを1つ示せばいいので、これで細胞株が違うことは証明できています。
2019/8/14(水) 午前 9:56[ カツラ報告書 ]返信する黒線のところが1Mb幅で半分のSNPs数の違っているところです。
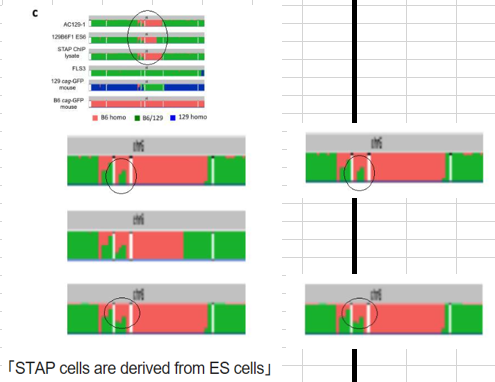
最初に指摘して置いたように、若山さんはややこしいことに取り下げ理由書にLetter Figure 4-bのSTAP細胞とES細胞のデータが取り違えられていると書いています。再掲します。
>>
(4) In Fig. 4b of the Letter, STAP cell and ES cell are wrongly labelled in a reverse manner.この取り違えは作表時なのか、サンプル提出時なのかわかりません。STAP ChIP lysateが提出時のES細胞の取り違えだったのなら、ここから129B6F1ESらしきものが出ても変ではありません。
ではその際は間違えられた方のESがSTAPだということになるが、そのマウス背景は何なのか。データ登録されたマウス背景はCD45がGOFで、TSがCD1、そしてその他は全部B6 X 129です。そしてそれを確かめるためのinputデータは揃っているが、検証はされていない。凍結lysateはSTAPだけだったのかどうかもわかってない。つまり、何事も系統立てては調べられていない。調査はとても恣意的で場当たり的だと言わざるを得ない。
- 2019/08/27(火) 09:34:47|
- AC129
-
-
| コメント:0
前のページ






































































